Introducción
Los estudios sobre la diversidad biológica de los individuos más tempranos de las Américas se remontan a casi 200 años atrás, cuando se encontraron y describieron los primeros esqueletos humanos de Sudamérica considerados antiguos (Lund, 1840, 1844; Ameghino, 1880; Hrdlička, Holmes, Willis, Wright y Fenner, 1912). Desde entonces, se han planteado varios interrogantes, tales como: dónde se originaron los ancestros de los individuos que llegaron a las Américas, cuándo llegaron, y cuáles fueron las dinámicas de expansión poblacional, por mencionar sólo algunos (Neves y Pucciarelli, 1989; Neves y Hubbe, 2005; Fagundes et al., 2008; González-José, Bortolini, Santos y Bonatto, 2008; Pucciarelli, Perez y Politis, 2010; de Azevedo et al., 2011; de Saint Pierre et al., 2012; Hubbe, Strauss, Hubbe y Neves, 2015; Raghavan et al., 2015; Skoglund et al., 2015; Posth et al., 2018; entre otros). Actualmente, gracias al aporte de distintas disciplinas, al creciente número de muestras arqueológicas disponibles, y a los avances tecnológicos de las últimas décadas, muchas de las preguntas pueden ser abordadas con mayor profundidad. Esto ha permitido avanzar en aspectos tales como el origen de la(s) población(es) ancestral(es) de las Américas, para el cual se considera que la mayor parte proviene del este de Asia (Moreno-Mayar et al., 2018a; Posth et al., 2018); y la antigüedad de los humanos en las Américas, que gracias al avance de estudios recientes se sostiene la presencia humana en el centro sur de Norteamérica antes del último máximo glacial (Ardelean et al., 2020; Boëda et al., 2021; Bennett et al., 2021; Madsen et al., 2022). Sin embargo, más allá de la gran atención que esta región ha recibido a lo largo de las últimas décadas, aún quedan muchos aspectos por ser estudiados e interpretados en mayor profundidad.
Uno de los aspectos que aún queda por dilucidar es aquel relacionado con las dinámicas de expansión de las poblaciones humanas a lo largo y a lo ancho de ambos continentes. Una vez llegados a Beringia, múltiples caminos podrían haber sido adoptados en la dispersión hacia el sur (Miotti, 2006; Achilli et al., 2008; Goebel, Waters y O’Rourke, 2008; Perego et al., 2010; Bodner et al., 2012; Reich et al., 2012; Achilli et al., 2013; Battaglia et al., 2013; Llamas et al., 2016). En Norteamérica, gracias al avance de estudios paleoambientales, arqueológicos y genéticos, existen dos hipótesis principales que explican la dispersión humana de esta amplia región: la ruta costera, a lo largo de la cual los primeros humanos se desplazaron a lo largo de la costa Pacífica y desde allí se expandieron tanto hacia al interior de Norteamérica como hacia Centroamérica y Sudamérica (Erlandson et al., 2007; Tamm et al., 2007; Fagundes et al., 2008; Bodner et al., 2012); y la ruta interior, según la cual se plantea que los desplazamientos se realizaron mediante el corredor libre de hielo entre las masas de hielo Laurentino y Cordillera (que cubrían lo que son actualmente las provincias de Alberta y el este de la Columbia Británica), cuya disponibilidad como pasaje se estima habría empezado hace 13.000 años AP (Pedersen et al., 2016; Potter et al., 2017). No se descarta la utilización de ambas rutas, lo cual inevitablemente queda supeditado a las condiciones climáticas que prevalecieron en diferentes momentos.
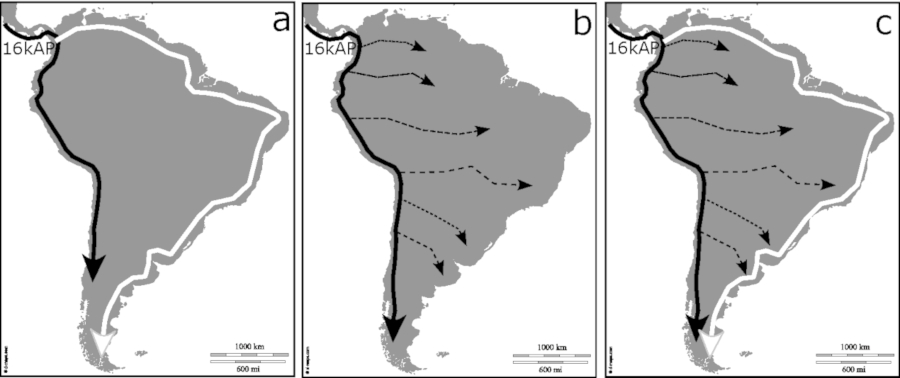
En base a expectativas arqueológicas y antropológicas previas, estudios genéticos recientes plantean que las expansiones humanas iniciales a Sudamérica habrían comenzado hace aproximadamente 16.000 años AP, luego de la divergencia entre los linajes que fueron descriptos como NNA/SNA (North Native Americans, South Native Americans), seguido de una expansión desde Mesoamérica que habría comenzado hace 8.000 años AP y que alcanzó Patagonia hace aproximadamente 5.000 años AP (Moreno-Mayar et al., 2018a; Posth et al., 2018). De acuerdo con interpretaciones de la evidencia arqueológica, lingüística, genética y osteológica, existen al menos dos modelos que explican la dispersión humana temprana en Sudamérica: a) una divergencia temprana en el noroeste de Sudamérica, a partir de la cual las poblaciones humanas se habrían bifurcado siguiendo, por un lado, una ruta andina a lo largo del océano Pacífico; y por otro lado, poblaciones más pequeñas habrían seguido un trayecto a lo largo de llanuras aluviales y/o a lo largo del océano Atlántico (Gruhn, 1994; Tarazona-Santos et al., 2001; Miotti, 2006; Pucciarelli et al., 2006; Wang et al., 2007; Rothhammer y Dillehay, 2009; Yang et al., 2010; Harris et al., 2018). La alternativa más fuerte a dicho modelo es b) una expansión inicial mediante una ruta costera a lo largo del Pacífico, a partir de la cual se habrían producido migraciones trasandinas hacia el este del continente (Wang et al., 2007; Bodner et al., 2012; Rothhammer, Fehren-Schmitz, Puddu y Capriles, 2017; Gómez-Carballa et al., 2018); aunque no se debería descartar c) una combinación de ambos modelos (Miotti, 2006; Menéndez, Rademaker y Harvati, 2019) (Figura 1). Luego, durante fines del Holoceno medio y comienzos del Holoceno tardío se estableció otra etapa de dinámicas expansivas que no serán abordadas en este trabajo, tales como el arribo de individuos de Mesoamérica desde hace aproximadamente 5.000 años (Moreno-Mayar et al., 2018a; Posth et al., 2018) y los movimientos de escala continental de grupos Tupí desde el Holoceno medio al tardío (Ramallo et al., 2013; Iriarte et al., 2017).
Figura 1
Modelos que explican la dispersión humana temprana en Sudamérica: (a) divergencia temprana en el noroeste de Sudamérica; (b) ruta costera a lo largo del Pacífico y migraciones trasandinas; (c) una combinación de los anteriores.
El análisis de la variación morfológica de los esqueletos humanos del Holoceno temprano y medio de las Américas puede contribuir al debate sobre las más antiguas dinámicas de expansión en el continente (Neves y Pucciarelli, 1989, 1991; Neves y Hubbe, 2005; Sardi, Rozzi, González-José y Pucciarelli, 2005; González-José et al., 2008; Perez, Bernal, Gonzalez, Sardi y Politis, 2009; Pucciarelli et al., 2010; de Azevedo et al., 2011; Manríquez et al., 2011; Hubbe et al., 2015; Menéndez et al., 2015; Delgado Burbano, 2017; von Cramon Taubadel, Strauss y Hubbe, 2017; Kuzminsky et al., 2018, entre otros). A partir del estudio de la variación craneométrica de los esqueletos humanos más tempranos del continente hallados a la fecha pueden discutirse posibles vías de dispersión con base en las similitudes y diferencias morfométricas entre individuos de distintas áreas (Pucciarelli et al., 2006, 2008). Se ha planteado previamente la existencia de una estructura geográfica este-oeste principalmente para poblaciones humanas recientes, donde las poblaciones del este fueron descritas presentando los niveles más bajos de variación intrapoblacional, neurocráneo de tamaño pequeño y grandes similitudes con los individuos más tempranos de Sudamérica (Tarazona-Santos et al., 2001; Pucciarelli et al., 2006). Asimismo, trabajos previos han demostrado que muchos de los individuos tempranos procedentes de distintas regiones presentan grandes similitudes entre sí, e.g., Sabana de Bogotá y Lagoa Santa (Neves et al., 2013; Hubbe et al., 2015), mientras que otros investigadores destacan la gran variación intrapoblacional e interpoblacional de las primeras poblaciones humanas que se dispersaron en las Américas (Gonzalez-José et al., 2008; Hubbe et al., 2020, entre otros). Sin embargo, aún quedan por analizar las causas de dicha variación morfológica, así como las similitudes y diferencias de individuos del Holoceno temprano/medio con el fin de interpretar dinámicas poblacionales de expansión.
El objetivo de este trabajo es analizar las afinidades morfológicas de algunos de los esqueletos más antiguos de Sudamérica con el fin de evaluar los modelos que describen patrones de expansión en el continente durante el Holoceno temprano y medio, es decir, momentos inmediatamente posteriores a las primeras expansiones humanas. Mediante el análisis de la variación morfológica los esqueletos humanos del Holoceno temprano y Holoceno medio inicial, discutiremos las dinámicas de expansión ocurridas previamente al cambio poblacional que tuvo lugar luego del Holoceno medio (“mid-Holocene population turnover”, sensuPosth et al., 2018). Pondremos a prueba el modelo según el cual se habría producido una divergencia temprana en el noroeste de Sudamérica, a partir de la cual las poblaciones humanas se habrían escindido siguiendo, ya sea una ruta andina o costera a lo largo del océano Pacífico y, por otro lado, un trayecto hacia el interior del continente y/o a lo largo del océano Atlántico. Si este modelo permite explicar la dispersión de las poblaciones humanas en Sudamérica, entonces esperamos encontrar una estructuración poblacional con muestras del oeste y este del continente que presenten las mayores diferencias entre sí, mientras que las muestras al interior de cada una de estas regiones deberían presentar mayores similitudes entre sí. Si, por el contrario, los resultados muestran similitudes entre individuos del oeste y este de Sudamérica, entonces dicho modelo no es suficiente para explicar las dispersiones humanas tempranas. Si este fuese el caso, el modelo que plantea una expansión inicial mediante una ruta costera a lo largo del Pacífico, a partir de la cual se habrían producido migraciones trasandinas hacia el este del continente, explicaría las primeras expansiones humanas en Sudamérica. Cabe destacar que aquí estamos evaluando el rango de ocupaciones inmediatamente posteriores a las expansiones más tempranas del continente, así como también los posibles movimientos ocurridos entre el Holoceno temprano y medio. Por lo tanto, no estamos considerando la existencia de migraciones más recientes ni de flujo génico entre regiones durante el Holoceno tardío, lo cual ha sido evaluado en detalle en otros trabajos, y dada su complejidad requiere un análisis pormenorizado entre las distintas regiones (García, Pauro, Nores, Bravi y Demarchi, 2012; Ramallo et al., 2013; Posth et al., 2018; Barberena et al., 2020; Postillone, Martínez, Flensborg y Dejean, 2020; entre otros).
Materiales y métodos
Muestra
Compilamos una muestra de 92 individuos adultos del Holoceno temprano (e.g., 11.700-8.200 años AP; Walker et al., 2012) y Holoceno medio (e.g., 8.200-4.200 años AP; Walker et al., 2012) procedentes del norte de Chile, Perú, pampas argentinas, Colombia, este de Brasil, México y Estados Unidos (Figura 2; Tabla 1). Los datos fueron obtenidos principalmente de la base de datos abierta recolectada por el Dr. Héctor Pucciarelli (Pucciarelli et al., 2006, 2008; Paschetta, González-José y Lanata, 2017), y la base de datos publicada por el Dr. Richard Jantz y el Dr. Douglas Owsley (Owsley y Jantz, 1999). Los nombres de las medidas (e.g., distancias lineales) registradas en ambos casos difieren dado que dichos investigadores tenían distintas preguntas de trabajo. Mientras que Jantz y Owsley estaban interesados en inferir la historia poblacional a partir de las muestras que estudiaron -para lo cual emplearon las variables definidas por W. W. Howells (Howells, 1995; Jantz y Owsley, 2001)-, Pucciarelli definió variables craneofuncionales, ya que su interés era evaluar la influencia de factores ambientales en la variación craneofacial (Pucciarelli, 2008; Sardi, 2017). Sin embargo, visto que, en algunos casos, las medidas morfométricas fueron definidas según los mismos puntos anatómicos, estas pueden considerarse homólogas a pesar de que tienen distintos nombres.
Tabla 1
Composición de las muestras utilizadas para el presente estudio e información cronológica asociada. La cronología se presenta de dos maneras: rangos cronológicos relativos basados en la estratigrafía indicados al comienzo con “~”, y fechados radiocarbónicos directos calibrados con un rango de dos sigmas indicados con “*”. En este último caso, se presenta el fechado entre paréntesis.
|
País/Región |
Sitio arqueológico |
Número de individuos |
Sexo (M/F/I) |
Cronología en años calibrados con dos sigma (cronología en años 14C AP) |
Referencia bibliográfica |
|
Estados Unidos |
Plattsmouth Oss, Nebraska |
2 |
1/1/0 |
~10,000-9,000 |
Owsley y Jantz (1999); Jantz y Owsley (2001) |
|
Wet Gravel Pit, Nebraska |
2 |
1/1/0 |
~10,000-9,000 |
Key (1982); Owsley y Jantz (1999); Jantz y Owsley (2001) |
|
Gilder Mound, Nebraska |
1 |
0/1/0 |
~10,000-9,000 |
Key (1982); Owsley y Jantz (1999); Jantz y Owsley (2001) |
|
Lime Creek, Nebraska |
1 |
1/0/0 |
~10,000-9,000 |
Key (1982); Owsley y Jantz (1999); Jantz y Owsley (2001) |
|
Buhl, Idaho |
1 |
0/1/0 |
12,772-12,474 (10,675 ± 95)* |
Green et al. (1998); Lepper (2014) |
|
Wizards Beach, Nevada |
1 |
1/0/0 |
10,516-10,245 (9,225 ± 60)* |
Tuohy y Dansie (1997); Owsley y Jantz (1999); Lepper (2014) |
|
Spirit Cave 2, Nevada |
1 |
1/0/0 |
10,720-10,571 (9,415 ± 25)* |
Tuohy y Dansie (1997); Jantz y Owsley (2001); Lepper (2014) |
|
México |
Peñón de los Baños III |
1 |
0/1/0 |
10,720-10,571 (10,755 ± 75)* |
Gonzalez et al. (2003, 2015); Pucciarelli, González-José, Neves, Sardi y Rozzi (2008) |
|
Tlapacoya 1 |
1 |
1/0/0 |
12,104-11,606 (10,200 ± 65)* |
Gonzalez et al. (2003, 2015); Pucciarelli, González-José, Neves, Sardi y Rozzi (2008) |
|
Metro Balderas |
1 |
1/0/0 |
~10,500 |
Pucciarelli González-José, Neves, Sardi y Rozzi (2008); Gonzalez et al. (2015) |
|
Paleoméxico Cuenca 10-613652 |
1 |
1/0/0 |
~10,000-9,000 |
Pucciarelli González-José, Neves, Sardi y Rozzi (2008) |
|
Chimalhuacán |
1 |
1/0/0 |
~10,500 |
Gonzalez et al. (2015); Pucciarelli González-José, Neves, Sardi y Rozzi (2008) |
|
Cueva del Tecolote |
1 |
1/0/0 |
~9,000-7,000 |
Gonzalez et al. (2015); Pucciarelli González-José, Neves, Sardi y Rozzi (2008) |
|
Colombia |
Checua |
9 |
7/2/0 |
9443-8307 (8,200 + 110 -6800 +)* |
Groot (1992); Correal Urrego y Ardila (1979); Pucciarelli et al. (2006); Delgado Burbano (2017) |
|
Tequendama |
10 |
6/4/0 |
8182-6480 (7269 + 65 - 5848 + 56)* |
Correal Urrego y van der Hammen (1977); Pucciarelli et al. (2006); Delgado Burbano (2015, 2017) |
|
Chile (Norte) |
Camarones 14 |
5 |
3/2/0 |
8724-6621(7420 + 225 - 6615 + 390)* |
Schiappacasse y Niemeyer (1984); Arriaza (1995); Pucciarelli et al. (2006) |
|
Perú |
Lauricocha |
4 |
3/1/0 |
8530-5748 (7871 + 30 -5158 + 27)* |
Cardich (1964); Pucciarelli et al. (2006); Fehren-Schmitz et al. (2015) |
|
Paiján |
1 |
1/0/0 |
11,945-8,452 (9,020 + 650)* |
Chauchat (1975); Chauchat y Lacombe (1984); Chauchat, Ivez, Briceno y Uceda (1998); Pucciarelli et al. (2006) |
|
Brasil (Este/Noreste) |
Serra da Capivara |
1 |
0/0/1 |
9526-9681 (8640 + 30)* |
Menéndez et al. (2022) |
|
Lagoa Santa |
39 |
27/12/0 |
~10000-7000 |
Pucciarelli et al. (2006); Da-Gloria, Neves y Hubbe (2017) |
|
Argentina (Pampas) |
La Tigra |
1 |
1/0/0 |
8177-7941 (7270 + 60)* |
Politis, Barrientos y Stafford (2011); Menéndez et al. (2015) |
|
Laguna de los Pampas |
1 |
1/0/0 |
10,238-9,762 (8971 + 77)* |
Politis, Messineo, González, Álvarez y Favier Dubois (2012); Menéndez et al. (2015) |
|
Necochea |
1 |
1/0/0 |
7938-7676 (7013 + 67)* |
Politis y Bonomo (2011); Menéndez et al. (2015) |
|
Arroyo del Moro |
2 |
1/1/0 |
7845-7571 (6885 + 73)* |
Politis y Bonomo (2011); Menéndez et al. (2015) |
|
Arroyo Seco 2 |
2 |
1/1/0 |
8554-8370 (7700 + 60)* |
Politis, Gutiérrez y Scabuzzo (2014); Menéndez et al. (2015) |
|
Arroyo Chocorí |
1 |
1/0/0 |
7882-7681 (7010 + 60)* |
Politis, Barrientos y Stafford (2011); Menéndez et al. (2015) |
|
TOTAL |
|
92 |
63/28/1 |
|
|
Figura 2
Mapa con la localización de las muestras analizadas.
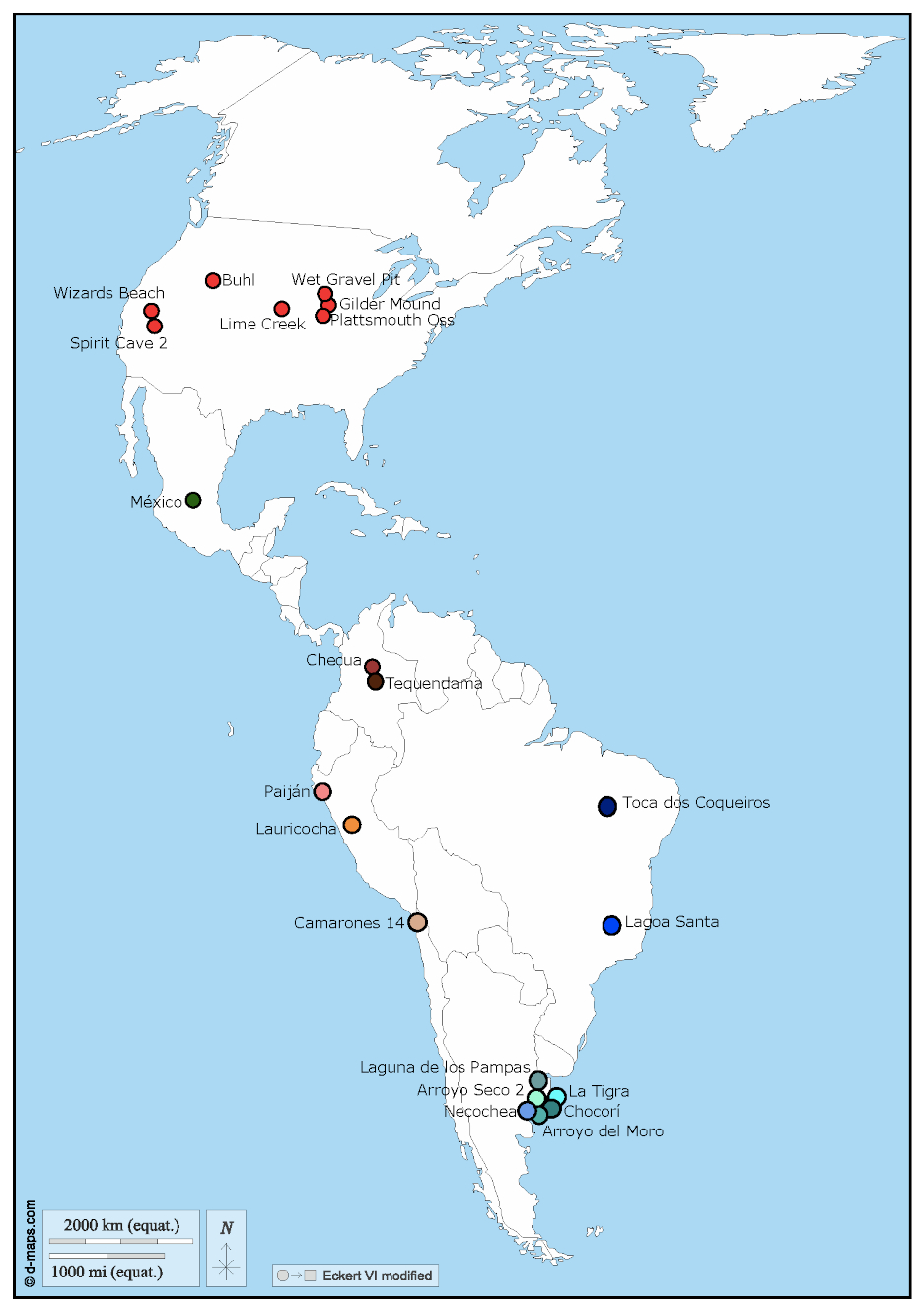
En la Tabla 1 se detalla el número de individuos, información cronológica asociada, y la estimación del sexo para cada una de las muestras. Mientras que algunas muestras presentan fechados radiocarbónicos directos, otras presentan información del contexto arqueológico que permite asociarlas a un rango cronológico. En el presente trabajo, las muestras fueron agrupadas por sitio arqueológico y también por región geográfica (correspondiente con el territorio político del país donde hoy se encuentran), siguiendo el agrupamiento utilizado previamente por Menéndez et al. (2015) para esta misma base de datos. Dado el carácter fragmentario de las muestras del Holoceno temprano, hemos decidido analizar todo el conjunto disponible, a pesar de que los distintos tamaños muestrales puedan afectar la interpretación de los resultados.
Variables morfométricas
Se seleccionó un total de diez medidas lineales que describen tanto rasgos morfométricos de la bóveda craneana como del esqueleto facial (Tabla 2). Estas diez medidas se encontraban presentes en ambas bases de datos morfométricas, y fueron registradas siguiendo a Pucciarelli (2008) y, en el caso de Jantz y Owsley (2001), siguiendo a Howells (1973). Las medidas fueron seleccionadas buscando encontrar un equilibro para maximizar tanto la inclusión de la mayor cantidad de individuos del Holoceno temprano en la muestra, como el número de variables para describir de manera más completa la variación morfológica de esta. Sin embargo, en muchos casos nos enfrentamos con dos dificultades: 1) los cráneos estudiados no se encuentran completos y, por ende, no permiten registrar un mayor número de medidas; 2) la cantidad de variables equivalentes entre ambas bases de datos es baja. De todas maneras, consideramos que las diez medidas lineales seleccionadas representan una solución intermedia cuando se compara con otros trabajos que utilizaron una muestra similar, algunos empleando solo seis variables (Steele y Powell, 1992), y otros, 22 variables (Jantz y Owsley, 2001).
Tabla 2
Lista de las medidas lineales craneofaciales usadas para este estudio.
|
Abreviatura |
Nombre de la medida (puntos craneométricos que la delimitan) |
|
LN |
Longitud neurocraneana (nasion-opistocraneo) |
|
AN |
Ancho neurocraneano (eurion-eurion) |
|
AF |
Ancho facial (cigion-cigion) |
|
HNM |
Altura neurocraneana media (basion-bregma) |
|
HF |
Altura facial (nasion-prostion) |
|
HR |
Altura respiratoria (nasion-nasoespinal) |
|
AR |
Ancho respiratorio (alar-alar) |
|
HO |
Altura óptica (punto supraorbitario medio-punto infraorbitario medio) |
|
AO |
Ancho óptico (dacrion-ectoconquio) |
|
AA |
Ancho alveolar (ectomolar-ectomolar) |
No hemos realizado análisis del error interobservador dado que no existen individuos en común en ambas bases de datos para los cuales se pueda calcular si existen diferencias en el registro de datos. Sin embargo, a pesar de que la base de datos de los individuos de Estados Unidos fue registrada por otros observadores distintos al resto de la muestra, consideramos relevante incluir tales muestras como comparativas en nuestro trabajo. Por esto es por lo que seguimos el criterio previamente aplicado por Delgado Burbano (2015), quien también utilizó dicha base de datos comparativa después de haber constatado que se siguieron los mismos procedimientos de registro de las variables aquí analizadas. Asimismo, a pesar de que las muestras no se encuentran balanceadas según el sexo de los individuos (63 individuos masculinos, 28 femeninos, 1 indeterminado), seguimos trabajos previos que han sido analizado individuos de esta base de datos sin ningún tipo de corrección preliminar por sexo (Pucciarelli et al., 2006, 2010; Menéndez et al., 2015, 2019). Esta base de datos contiene la mayoría de los individuos más antiguos recuperados en las Américas, por lo que dicho sesgo se encuentra asociado con la visibilidad arqueológica en relación con los entierros del Pleistoceno final y del Holoceno temprano (Dillehay, 1997).
Las distancias lineales que fueron registradas por los observadores de ambas bases de datos constituyen variables de forma (form). La forma (form en inglés) está compuesta por forma (shape en inglés) y tamaño (centroid size) (Zelditch, Swiderski, Sheets y Fink, 2004). Dado que el objetivo de este trabajo es analizar la variación en forma (shape), calculamos las razones de Mosimann para poder obtener variables que excluyan la información de tamaño geométrico (i.e., media geométrica). Estas razones fueron calculadas como el cociente entre cada una de las distancias y la media geométrica de la totalidad de las distancias por cada individuo (Darroch y Mosimann, 1985; Jungers, Falsetti y Wall, 1995). Todos los siguientes análisis fueron realizados empleando las variables de forma (shape).
Análisis estadísticos
La variación morfológica de los individuos de las Américas fue explorada por medio de un análisis de componentes principales (ACP). Este método es ampliamente utilizado en estudios morfométricos, ya que permite reducir la dimensionalidad de los datos estudiados, al mismo tiempo que resume y ordena el porcentaje de la variación descrita por las diferentes variables (Mitteroecker y Gunz, 2009). Mediante el ACP se transforma el conjunto original de variables (aquí, las diez medidas craneométricas) en un nuevo conjunto de variables no correlacionadas entre sí que representan la mayor variación del conjunto de datos (Zar, 2010). Se realizó un ACP utilizando la totalidad de los individuos y agrupando las muestras por región geográfica, lo cual se corresponde con el actual territorio político de cada país (ver Tabla 1). Complementariamente, utilizando los scores de los diez componentes principales (CP) que resumen la variación total en la muestra se realizó un análisis de funciones discriminantes (AFD). El AFD es un análisis estadístico multivariado que se diferencia del ACP porque su propósito es encontrar un eje de combinación de las variables que permita maximizar la diferencia existente entre grupos definidos a priori (Zar, 2010). Asimismo, el AFD estima la distancia que hay entre cada individuo y el centroide de cada grupo usando distancias Mahalanobis, que permiten clasificar las muestras según las variables dentro de los grupos previamente establecidos. Ya que las muestras son asignadas al grupo más cercano, dicha clasificación nos permite evaluar afinidades entre individuos. Tomando en cuenta la heterogeneidad que existe en el tamaño muestral, llevamos a cabo el AFD especificando las probabilidades a priori para cada grupo proporcional a la frecuencia de individuos por grupo. De esta manera, presentamos los gráficos que muestran la dispersión de las muestras a lo largo de los dos primeros componentes principales, y de los dos primeros ejes discriminantes, así como también tablas que muestran la contribución de las variables utilizadas en los dos primeros componentes y ejes principales. Complementariamente, reportamos la matriz de la tasa de clasificación predicha para cada grupo, la cual resume las probabilidades posteriores esperadas para cada muestra.
Las afinidades biológicas entre los individuos tempranos de las Américas fueron evaluadas mediante distancias biológicas. Para esto calculamos distancias Mahalanobis, mismas que son ampliamente utilizadas para evaluar similitudes entre poblaciones, ya que consideran la posible correlación entre las variables aleatorias, además de no verse afectadas por la escala y la unidad de las variables (Mahalanobis, 1930). Las distancias Mahalanobis fueron representadas mediante un análisis de coordenadas principales (ACoP), cuyos resultados se presentan mediante un gráfico de dispersión de los dos primeros ejes (Davis, 1986). Para el cálculo de distancias y su representación visual, se agruparon las muestras por sitio arqueológico usando la media de las medidas morfométricas de los individuos agrupados según el sitio arqueológico, aunque para México y Estados Unidos utilizamos las medias del conjunto. Dado que el presente estudio apunta a discutir dinámicas de expansión en Sudamérica, las muestras de México y Estados Unidos son utilizadas como comparativas, y por eso decidimos mantener a los individuos agrupados en estas dos regiones.
El análisis de componentes principales, el análisis de coordenadas principales y el análisis discriminante fueron realizados con los programas PAST (v. 2.17, Hammer, Harper y Ryan, 2001) y JMP (v. 16, by the SAS Institute Inc., 2021), mientras que las distancias Mahalanobis fueron calculadas en R (R Core Team, 2021).
Resultados
Variación morfológica de los individuos tempranos de las Américas
El ACP realizado con la totalidad de los individuos agrupados por región geográfica muestra que los dos primeros componentes principales (CP) explican el 50% de la variación total de la muestra (Figura 3A). A pesar de que existe una gran superposición en el rango de variación, al observar la distribución de las muestras a lo largo del CP2 se puede ver que los individuos del este/noreste de Sudamérica (Brasil) se encuentran hacia el extremo positivo, mientras que los individuos provenientes de la región andina o el oeste del continente (Colombia, Perú, Chile) se encuentran hacia el extremo negativo de dicho rango (Figura 3A). Adicionalmente, las muestras de Norteamérica (México, Estados Unidos) se ubican hacia el centro de la distribución, aunque más cercanas a las del oeste de Sudamérica, mientras que las de Argentina se distribuyen a lo largo de toda la variación del CP2, y ocupan un lugar central en la distribución. El CP1 no muestra diferencias que puedan explicarse según los criterios aquí seguidos; otros factores que exceden a los objetivos de este trabajo contribuirían a explicar la variación a lo largo de este componente. Sin embargo, se destaca que algunas muestras presentan una amplia variación a lo largo del CP2 (Brasil, Colombia, Perú), mientras que otras presentan una variación más acotada (Argentina, Chile). Las muestras de Norteamérica (México, Estados Unidos) se encuentran hacia el extremo negativo del CP1, presentan un rango acotado de variación y una fuerte superposición. La Tabla 3 muestra la contribución que tiene cada una de las variables morfométricas en los componentes principales. Para el CP1, puede verse que las variables que mayormente contribuyen son el ancho facial (AF) y la altura neurocraneana media (HNM). Los individuos del este de Sudamérica (Brasil, y parte de los individuos de las pampas argentinas) presentan esqueletos faciales más angostos que los individuos del noroeste del continente (Colombia, Perú). Complementariamente, aunque en menor medida, los individuos del noroeste del continente presentan bóvedas craneanas más bajas en sentido superior-inferior, mientras que, en comparación, los individuos de Brasil, y parte de los de las pampas presentan bóvedas más altas.
Figura 3
Análisis de la variación morfológica de los individuos tempranos de las Américas, agrupados por región geográfica: A) Dispersión de las muestras a lo largo de los dos primeros componentes principales; B) Dispersión de las muestras a lo largo de los dos primeros ejes discriminantes.
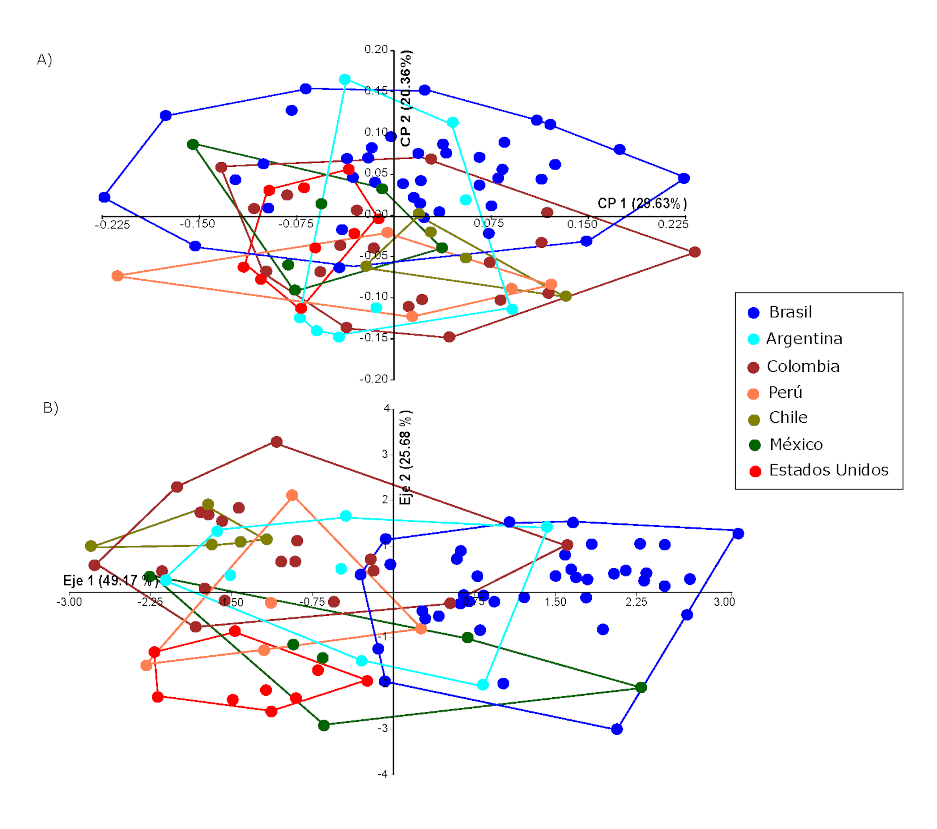
Tabla 3
Contribución de las variables utilizadas para este estudio en cada componente principal (CP) o eje discriminante (ED); las dos primeras columnas refieren al ACP de todos los individuos agrupados por región; las columnas tercera y cuarta, al AD de todos los individuos agrupados por región. En negrita se indican las variables que mayormente contribuyen a maximizar la variación de la muestra (ACP) o de los grupos (AD).
|
|
PCA todos los individuos por región |
Análisis discriminante todos los individuos por región |
|
Componente o eje Principal |
1 |
2 |
1 |
2 |
|
LN |
0.256 |
0.047 |
4.020 |
-0.561 |
|
AN |
-0.111 |
0.234 |
-3.804 |
1.523 |
|
AF |
-0.847 |
0.082 |
-0.914 |
-3.147 |
|
HNM |
0.400 |
0.515 |
1.585 |
-0.924 |
|
HF |
0.091 |
-0.757 |
-21.675 |
-9.869 |
|
HR |
0.175 |
-0.293 |
8.071 |
22.379 |
|
AR |
0.032 |
0.012 |
2.638 |
9.856 |
|
HO |
0.047 |
0.002 |
-3568 |
7.995 |
|
AO |
-0.003 |
0.091 |
0.666 |
-17.330 |
|
AA |
-0.041 |
0.066 |
12.980 |
-9.923 |
La variación a lo largo de los dos primeros ejes discriminantes (ED) resume un 75% de la variación de la muestra, y presenta una moderada estructuración geográfica (Figura 3B). El ED1 diferencia las muestras de Sudamérica en sentido oeste-este. Las muestras de Colombia, Chile, y Perú se encuentran en el extremo negativo del ED2, mientras que, en el extremo positivo, se ubican las de Brasil (Figura 3B). Las variables que mayormente contribuyen a las diferencias a lo largo del ED1 son la altura facial, altura respiratoria, y ancho alveolar (Tabla 3). Las muestras de Sudamérica, a diferencia de las de Norteamérica, presentan esqueletos faciales más altos en sentido supero-inferior, maxilares más amplios y narices más altas. El ED2 separa a las muestras en sentido norte-sur; las muestras de Norteamérica (Estados Unidos, México) se ubican hacia el extremo negativo, mientras las muestras de Sudamérica se encuentran hacia el extremo positivo (Colombia, Chile, Perú). Algunos individuos de Perú, Argentina, y Brasil presentan afinidad con las muestras de Norteamérica. Las muestras de Argentina se encuentran en una posición central considerando ambos EDs; algunos individuos son similares a aquellos de Brasil y otros a las muestras del oeste del continente (Figura 3B). Algunos individuos de Colombia y México se encuentran más cercanos a la distribución de Brasil. Las variables que mayormente contribuyen a dicha diferenciación son la altura respiratoria y la altura óptica, de manera que las muestras del oeste del continente presentan narices y órbitas más bajas que las de los individuos de Brasil (Tabla 3). La matriz de clasificación muestra que los grupos de Brasil, Colombia, Chile y Estados Unidos son los más homogéneos; los dos últimos son los que muestran tasas de clasificación correcta más altas (1.000 y 0.889, respectivamente). En contraste, las muestras de México, Perú, y Argentina presentan una mayor variación intramuestral, con valores entre 0.667 y 0.375 (Tabla 4). Esto confirma los resultados previos, de acuerdo con los cuales algunos individuos de Argentina presentan grandes afinidades con individuos de Colombia y Brasil, mientras que las muestras de México presentan afinidades morfológicas con las de Brasil.
Tabla 4
Matriz clasificatoria del análisis discriminante. Resumen de la tasa de clasificación predicha para cada grupo. Se destacan en negrita los valores mencionados en el texto.
|
Real |
Tasa predicha |
|
|
Argentina |
Brasil |
Chile |
Colombia |
EUA |
México |
Perú |
|
Argentina |
0.375 |
0.250 |
0.125 |
0.250 |
0.000 |
0.000 |
0.000 |
|
Brasil |
0.025 |
0.875 |
0.000 |
0.000 |
0.025 |
0.050 |
0.025 |
|
Chile |
0.000 |
0.000 |
1.000 |
0.000 |
0.000 |
0.000 |
0.000 |
|
Colombia |
0.053 |
0.105 |
0.000 |
0.842 |
0.000 |
0.000 |
0.000 |
|
EEUU |
0.000 |
0.000 |
0.000 |
0.000 |
0.889 |
0.111 |
0.000 |
|
México |
0.000 |
0.167 |
0.000 |
0.167 |
0.000 |
0.667 |
0.000 |
|
Perú |
0.000 |
0.200 |
0.000 |
0.200 |
0.200 |
0.000 |
0.400 |
Cabe destacar que las interpretaciones de los resultados del análisis de componentes principales y análisis discriminante pueden verse afectadas por el tamaño muestral diferencial.
Afinidades biológicas entre los individuos tempranos de las Américas
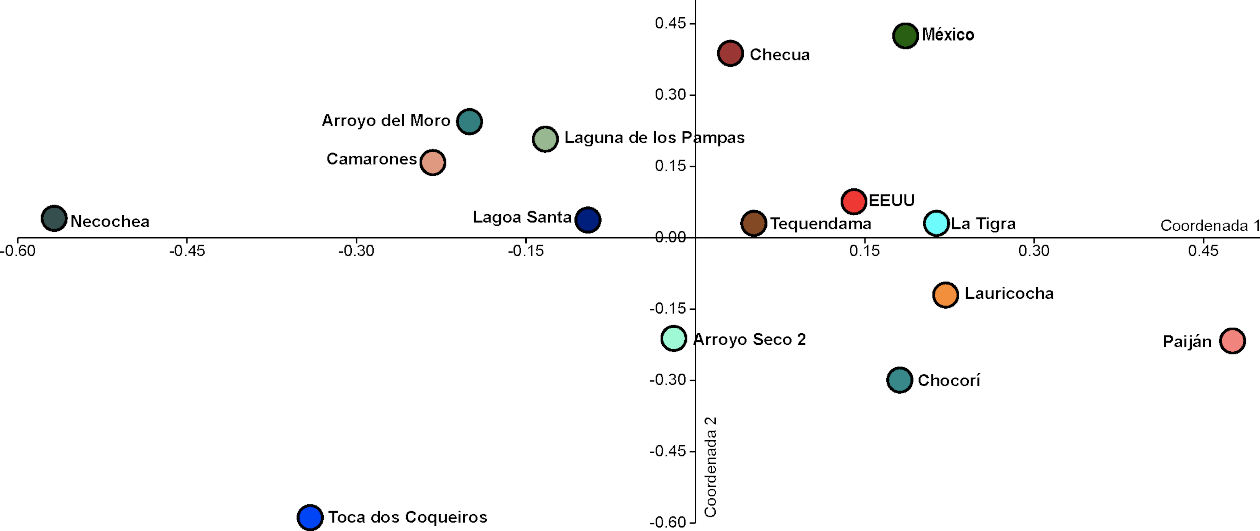
Los resultados de la evaluación de las afinidades biológicas entre individuos procedentes de sitios arqueológicos del Holoceno temprano y medio muestran que existe cierta asociación entre algunas muestras en función de cercanía geográfica, cronología, o las rutas de dispersión utilizadas (Tabla 5). Las distancias Mahalanobis permiten vislumbrar cierto patrón geográfico y cronológico. Los individuos más similares entre sí son aquellos de Arroyo Seco 2/Tequendama, Camarones/Arroyo del Moro, Lagoa Santa/Arroyo Seco 2, Tequendama/Checua; Tequendama/Camarones; Estados Unidos/México; Lagoa Santa/Tequendama; Tequendama/Arroyo del Moro, y Lauricocha/Estados Unidos (Tabla 5). Mientras que en la mayoría de los casos estas muestras presentan distancias geográficas medias (salvo Checua/Tequendama, que son muy próximas entre sí), las muestras de Lagoa Santa/Arroyo Seco 2 se encuentran en el este de Sudamérica; las de Checua/Tequendama, Camarones/Tequendama en el oeste del continente; y las muestras de Camarones/Arroyo del Moro se encuentran ambas en el Cono Sur (Tabla 5). No existen similitudes entre muestras muy distantes geográficamente, como podría ser Estados Unidos/Arroyo Seco 2. Asimismo, la muestra de Camarones/Arroyo del Moro y Tequendama/Arroyo del Moro presenta una superposición cronológica ~7800-7500 años AP, mientras Camarones/Tequendama presentan una superposición de ~8100-6600 años AP (Tabla 1); sin embargo, es interesante observar que, a pesar de que la muestra de Tequendama presenta similitudes con la de Arroyo Seco 2, estas no presentan un rango cronológico similar (Tequendama es más reciente). Asimismo, Lagoa Santa/Arroyo Seco 2 presentan una superposición cronológica alrededor de ~8500-8300 años AP (Tabla 1) y ambas se encuentran ubicadas en el este del continente, pero los individuos no parecen presentar similitudes morfológicas. Una gran cantidad de pares de muestras presentan diferencias significativas entre sí. Entre las muestras que más difieren entre sí se pueden destacar las de Laguna de los Pampas y Paiján, que presentan grandes diferencias con la mayoría de las otras. La muestra de Laguna de los Pampas presenta grandes diferencias con otras de la misma región: Necochea, Chocorí, Arroyo del Moro, Arroyo Seco 2, pero también con Camarones y Paiján. La muestra de Paiján presenta grandes diferencias con las muestras de Toca dos Coqueiros y Estados Unidos (Tabla 5). Dichas muestras no presentan superposiciones en los rangos cronológicos; algunas son del Holoceno medio, y otras, del temprano (Tabla 1). A lo largo de la primera coordenada del PCoA se puede ver cierta estructuración geográfica de acuerdo con la cual las muestras del oeste del continente (Paiján, Lauricocha, Tequendama, Checua) y las de Norteamérica (Estados Unidos, México) se encuentran hacia el extremo positivo, y las del este, hacia el extremo negativo (Laguna de los Pampas, Arroyo Seco 2, Lagoa Santa, Arroyo del Moro, Necochea, Toca dos Coqueiros), aunque existen algunas excepciones a este patrón (Camarones, La Tigra, Chocorí) (Figura 4). Particularmente, Paiján, Toca dos Coqueiros y Necochea son las más disímiles de toda la muestra, y se ubican en los extremos de la distribución (Figura 4).
Tabla 5
Distancias Mahalanobis (D2) de los individuos según el sitio arqueológico o región. Las D2 se indican por debajo de la diagonal, mientras que los valores de p asociados se encuentran por encima de ella. EEUU=Estados Unidos; MX=México; CAM=Camarones 14; CHE=Checua; LAU=Lauricocha; COQ=Toca dos Coqueiros; LS=Lagoa Santa; LT=La Tigra; LLP=Laguna de los Pampas; NE=Necochea; AM=Arroyo del Moro; AS=Arroyo Seco 2; CHO=Chocorí; PJ=Paiján; TQ=Tequendama. Se destacan en negrita los valores de distancias más altos y más bajos que se describen en el texto.
|
|
EEUU |
MX |
CAM |
CHE |
LAU |
COQ |
LS |
LT |
LLP |
NE |
AM |
AS |
CHO |
PJ |
TQ |
|
EEUU |
0 |
0.862 |
0.028 |
0.143 |
0.753 |
0.572 |
0.196 |
0.002 |
0.000 |
0.000 |
0.004 |
0.064 |
0.000 |
0.000 |
0.222 |
|
MX |
05.40 |
0 |
0.080 |
0.165 |
0.554 |
0.118 |
0.709 |
0.015 |
0.000 |
0.000 |
0.050 |
0.364 |
0.009 |
0.000 |
0.667 |
|
CAM |
20.03 |
16.71 |
0 |
0.295 |
0.039 |
0.000 |
0.100 |
0.000 |
0.000 |
0.000 |
0.945 |
0.456 |
0.110 |
0.000 |
0.825 |
|
CHE |
14.70 |
14.16 |
11.84 |
0 |
0.578 |
0.032 |
0.264 |
0.000 |
0.000 |
0.000 |
0.319 |
0.556 |
0.000 |
0.000 |
0.861 |
|
LAU |
06.70 |
08.76 |
19.07 |
8.52 |
0 |
0.113 |
0.283 |
0.000 |
0.000 |
0.002 |
0.083 |
0.222 |
0.000 |
0.000 |
0.553 |
|
COQ |
08.57 |
15.39 |
36.59 |
19.71 |
15.52 |
0 |
0.070 |
0.000 |
0.002 |
0.000 |
0.000 |
0.022 |
0.000 |
0.000 |
0.006 |
|
LS |
13.51 |
7.16 |
15.98 |
12.31 |
12.02 |
17.17 |
0 |
0.001 |
0.000 |
0.000 |
0.143 |
0.977 |
0.000 |
0.000 |
0.772 |
|
LT |
26.79 |
21.87 |
37.04 |
44.35 |
32.28 |
45.45 |
29.29 |
0 |
0.000 |
0.000 |
0.000 |
0.000 |
0.000 |
0.000 |
0.000 |
|
LLP |
32.76 |
40.20 |
72.32 |
50.15 |
53.03 |
27.32 |
41.40 |
61.10 |
0 |
0.000 |
0.000 |
0.000 |
0.000 |
0.000 |
0.000 |
|
NE |
41.91 |
30.03 |
30.92 |
32.51 |
26.78 |
74.78 |
40.57 |
45.71 |
105.93 |
0 |
0.046 |
0.000 |
0.000 |
0.000 |
0.005 |
|
AM |
25.39 |
18.27 |
4.04 |
11.49 |
16.59 |
43.48 |
14.68 |
37.81 |
78.39 |
18.53 |
0 |
0.384 |
0.026 |
0.001 |
0.773 |
|
AS |
17.46 |
10.91 |
9.81 |
8.74 |
13.02 |
20.75 |
3.16 |
35.53 |
56.51 |
40.83 |
10.65 |
0 |
0.011 |
0.000 |
0.973 |
|
CHO |
37.43 |
23.46 |
15.63 |
35.82 |
38.31 |
56.52 |
30.86 |
30.58 |
96.79 |
38.90 |
20.26 |
22.76 |
0 |
0.000 |
0.035 |
|
PJ |
62.67 |
47.98 |
36.35 |
34.83 |
46.18 |
65.01 |
43.60 |
79.95 |
106.96 |
51.49 |
28.12 |
32.37 |
34.83 |
0 |
0.000 |
|
TQ |
13.01 |
7.59 |
5.87 |
5.41 |
8.77 |
24.49 |
6.48 |
31.89 |
56.62 |
24.95 |
6.47 |
3.30 |
19.39 |
33.48 |
0 |
Figura 4
Análisis de coordenadas principales que muestra las afinidades biológicas entre los individuos de las Américas en base a distancias Mahalanobis entre las muestras a lo largo de las dos primeras dimensiones.
Discusión
En este artículo se analizó la variación morfológica de un conjunto de individuos del Holoceno temprano y medio de las Américas, con el fin de evaluar las afinidades biológicas entre ellos para discutir posibles modos de expansión de los descendientes de los primeros humanos en Sudamérica. El linaje de grupos Nativos Sudamericanos (SNA: South Native Americans) divergió hace ~16,000 años AP, lo cual coincide con estimaciones de la llegada de los primeros humanos a Sudamérica basadas en la distribución de fechados directos calibrados (Prates, Politis y Perez, 2020; Moreno-Mayar et al., 2018b). Sin embargo, cabe destacar que, actualmente, el esqueleto más antiguo de Sudamérica reportado con fechado radiocarbónico directo proviene de la Cueva de Candonga, en Córdoba, Argentina (media 2 sigma: 12.272 años calendáricos AP; Cornero, Neves y Rivero, 2014), lo que significa que los esqueletos del Holoceno temprano que han sido hasta ahora recuperados -muchos de los cuales forman parte de las bases de datos que fueron aquí analizadas- no representan los descendientes directos de los primeros migrantes, ya que son ~200 generaciones más recientes. Es decir, los esqueletos más antiguos disponibles para ser estudiados pertenecen a individuos que ya estaban asentados en Sudamérica, por lo que su variación morfológica brinda información acerca de posibles dinámicas de expansión en el continente durante los primeros milenios. En este trabajo en particular, nos focalizamos en la variación morfológica de los individuos recuperados que vivieron durante el Holoceno temprano y medio.
Nuestros resultados muestran que existe una relativa estructuración geográfica este-oeste de las muestras estudiadas, lo cual en parte sustenta el modelo de divergencia poblacional temprana en el noroeste de Sudamérica como resultado de una separación en al menos dos grupos que siguieron distintas direcciones (Gruhn, 1994; Tarazona-Santos et al., 2001; Miotti, 2006; Pucciarelli et al., 2006; Wang et al., 2007; Rothhammer y Dillehay, 2009; Yang et al., 2010; Harris et al., 2018). Las similitudes morfológicas entre individuos del este de Sudamérica (Brasil, Argentina) y sus diferencias con muestras del oeste del continente (Perú, Colombia) -lo cual ha sido ilustrado con los resultados del ADF, ACP, y PCoA- provee sustento al modelo según el cual los primeros grupos humanos que llegaron al noroeste de Sudamérica se separaron en dos grupos que migraron de manera independiente, uno siguiendo la costa Pacífica y el otro siguiendo la costa Atlántica o el interior del continente. Es decir, la diversificación morfológica basal entre poblaciones del este y oeste de Sudamérica se remontaría a ~16,000 años AP (Moreno-Mayar et al., 2018a; Prates et al., 2020). En este trabajo hemos asumido la existencia de un pulso de expansión antes del Holoceno medio (Rothhammer y Dillehay, 2009; Moreno-Mayar et al., 2018a; Posth et al., 2018), pero no debe descartarse que la descripta diferenciación entre muestras del este y oeste del continente puede reflejar una contribución diferencial de varios linajes ancestrales (Pucciarelli et al., 2006). Asimismo, la gran variación morfológica de la muestra -fundamentalmente evidenciada a través de los individuos de Argentina- apoya trabajos previos que plantean que las poblaciones del este de Sudamérica presentan mayor variación intragrupal que las poblaciones del oeste del continente (Tarazona-Santos et al., 2001; Pucciarelli et al., 2006; Menéndez et al., 2015), y trabajos que proponen una diversidad morfológica alta para los individuos del Holoceno temprano de las Américas (Sardi et al., 2005; González-José et al., 2008; Hubbe et al., 2020) y particularmente para Brasil (Menéndez et al., 2022). Sin embargo, esta afirmación no se mantiene en todos los casos ya que las muestras de Brasil se caracterizan por una gran homogeneidad intrapoblacional, lo cual concuerda con lo descripto anteriormente (Hubbe et al., 2015).
La amplia variación morfológica que muestran los individuos provenientes de las pampas argentinas, las similitudes que presentan con individuos de áreas geográficas distantes e incluso del oeste del continente (Camarones, Tequendama), así como las grandes diferencias entre individuos de esta región (Laguna de los Pampas con Necochea, Arroyo del Moro, Chocorí, Arroyo Seco 2) complejiza la interpretación de dicho modelo. Es decir, si bien el modelo de divergencia temprana en el noroeste de Sudamérica explica la variación de la mayoría de los individuos analizados, no explica completamente la variación morfológica de los individuos del Holoceno temprano y medio aquí analizados. La gran variación que presentan las muestras de las pampas argentinas, ilustradas por los resultados del ACP y el PCoA donde algunos individuos muestran mayores afinidades entre sí (Arroyo del Moro/Arroyo Seco 2), otros con individuos de Brasil (Arroyo Seco 2/Lagoa Santa), mientras que otros con muestras de Chile (Arroyo del Moro/Camarones), apoya la hipótesis de que algunas poblaciones del Cono Sur derivan de dos linajes que se habrían expandido independientemente, uno Pacífico/Andino y otro Amazónico/Atlántico (sensuGómez-Carballa et al., 2018). Estos resultados coinciden con resultados craneométricos previos que muestran una gran variación morfológica en individuos del Holoceno temprano de las pampas argentinas, y su asociación con individuos tempranos de otras regiones distantes tales como Lagoa Santa (Pucciarelli et al., 2010; Menéndez et al., 2015, 2017), y estudios genéticos que muestran flujo génico entre individuos de las pampas y de los Andes Centrales (Muzzio et al., 2018). La variación morfológica que ilustran los resultados del ACP y ADF, donde las muestras de Pampa se encuentran en el centro de la distribución, y las muestras de Brasil y Colombia en los extremos, apoyan esta interpretación. Sin embargo, debe tenerse en cuenta que esta puede estar influenciada por el bajo tamaño muestral de la muestra aquí analizada de las pampas argentinas, que no permite poner a prueba estadísticamente dicha hipótesis ni llegar a interpretaciones conclusivas. Asimismo, otro factor que también debe tenerse en cuenta cuando se analizan los cambios en la morfología craneofacial es el impacto de factores no-aleatorios, los cuales aquí no fueron evaluados, pero ya han sido planteados previamente en muestras del Cono Sur de Sudamérica (Perez y Monteiro, 2009; Fabra y Demarchi, 2011; Perez et al., 2011; Menéndez, 2018).
Asimismo, las grandes diferencias morfológicas que encontramos entre el individuo de Laguna de los Pampas respecto de otros de la misma región pueden explicarse como resultado de que aquel representa un linaje basal aún más antiguo, mientras los otros individuos representan linajes más recientes o mestizajes entre unos y otros (Roca-Rada et al., 2021). El fechado más antiguo que presenta este sitio (es el cráneo más antiguo con fechado directo en Sudamérica), en comparación con los otros, provee sustento a dicha interpretación (media 2 sigma: 9997 años calendáricos AP; Politis et al., 2012). Complementariamente, nuestros resultados muestran que el individuo de Arroyo Seco 2 se encuentra en el centro de la distribución en cuanto a la variación morfológica de las muestras de las pampas argentinas, lo cual apoya estudios previos que describen a las muestras de Arroyo Seco 2 como mixtas con un componente de Lagoa Santa/Anzick, pero también con un componente común a las otras poblaciones sudamericanas del Holoceno medio (Posth et al., 2018; Roca-Rada et al., 2021). Esto sugiere que durante el Pleistoceno final al Holoceno medio habrían confluido en las pampas argentinas poblaciones provenientes de linajes independientes que habrían llegado a través del Atlántico, pero también poblaciones que habrían migrado como producto de expansiones trasandinas. Es decir, consideramos que en las pampas argentinas habría existido continuidad poblacional durante el Holoceno temprano y medio con un moderado influjo de individuos asociados a linajes diferentes (Politis, 2008; Messineo et al., 2019; Postillone et al., 2020). De esta manera, los resultados de nuestro trabajo podrían interpretarse según un modelo de expansiones de linajes ancestrales de distintas procedencias (Figura 1c) para el caso de las pampas argentinas.
Con relación a la variación de las muestras del oeste de Sudamérica, las grandes similitudes entre algunas muestras (Checua/Paiján, Tequendama/Lauricocha), y también con las muestras de Norteamérica (Estados Unidos/Lauricocha, Estados Unidos/Tequendama), puede deberse a que estas derivarían de un mismo linaje ancestral. Esto concuerda con trabajos previos, en los cuales, a partir de análisis integrales de datos genéticos y morfológicos, se planteó que la falta de diferenciación genética y la moderada diversificación morfológica de muestras de la sabana de Bogotá puede deberse a un proceso de asimilación entre poblaciones locales y poblaciones de regiones adyacentes (Delgado Burbano, Rodríguez, Kassadjikova y Fehren-Schmitz, 2021). Además, estudios genéticos muestran una asociación estrecha entre muestras de Sudamérica y el individuo de Anzick-1 del noroeste de Estados Unidos (Posth et al., 2018), lo cual indica que el individuo de Anzick-1 formaba parte del mismo linaje ancestral del cual divergieron las poblaciones de Sudamérica. Respecto del centro y sur de los Andes, en Perú, las moderadas diferencias que encontramos entre los individuos de Lauricocha y el individuo de Paiján pueden explicarse por el relativo aislamiento geográfico de los grupos humanos que vivían en la sierra y en la costa. Según estudios genéticos recientes, dicha diversificación entre poblaciones de la sierra y de la costa tiene una profundidad temporal que se remonta a, al menos, ocho milenios (Lindo et al., 2018; Harris et al., 2018; Nakatsuka et al., 2020). Las similitudes entre las muestras del oeste de Sudamérica pueden interpretarse como resultado de la expansión a lo largo de la costa del Pacífico (Kuzminsky, 2013; Kuzminsky et al., 2018), mientras que las similitudes de la muestra de Chile con individuos de Pampa (Arroyo del Moro/Camarones) apoyan la existencia de migraciones trasandinas que se describieron más arriba, desde la región del Pacífico hacia el interior del continente (Harris et al., 2018; Gómez-Carballa et al., 2018).
Finalmente, las muestras de Norteamérica presentan una variación morfológica moderada, con fuertes afinidades y una gran superposición entre sí. Además, en general, se encuentran mayormente asociadas a las muestras del oeste de Sudamérica. Creemos que este patrón de afinidades podría explicarse como resultado de la expansión ocurrida de Mesoamérica hacia Sudamérica durante el Holoceno medio (Moreno-Mayar et al., 2018a). Asimismo, la ausencia de la señal Austromelanesia entre los individuos del oeste de Sudamérica (de acuerdo con Moreno-Mayar et al., 2018a, por el momento únicamente presente en individuos de Lagoa Santa; aunque según Posth et al., 2018, probablemente atribuible a un sesgo metodológico en los análisis realizados por Moreno-Mayar et al., 2018a), y la divergencia temprana de los individuos del oeste de Sudamérica (Harris et al., 2018) podrían explicar la menor variación de los individuos de Norteamérica y del oeste de Sudamérica. Sin embargo, estas muestras son de tamaño pequeño y han sido agrupadas para fines comparativos, por lo que se espera que el análisis de la variación de los individuos de Norteamérica pueda explorarse en más detalle en futuros trabajos.
Para concluir, nuestros resultados apoyan que la morfología craneofacial es una aproximación útil para discutir dinámicas de dispersión humana en Sudamérica. Asimismo, muestran que la dispersión humana en Sudamérica es más compleja que lo que ha sido planteado en los recientes modelos. Si bien el modelo de divergencia temprana en el noroeste de Sudamérica explica gran parte de la estructuración geográfica entre muestras del este y oeste del continente, hay ciertas regiones como las pampas argentinas para las cuales dicho modelo no resulta válido. Cabe destacar que para esta región existe una gran cantidad de sitios arqueológicos con pocos individuos por sitio (a excepción de Arroyo Seco 2, para el cual, de todas maneras, analizamos un individuo solamente). Para las otras regiones (e.g., Lagoa Santa, Brasil; Tequendama, Colombia), existe una gran cantidad de individuos provenientes de un mismo sitio arqueológico. Es decir, la gran variación que encontramos en la muestra de las pampas argentinas puede deberse a que no contamos con una representación de una gran cantidad de individuos de un solo sitio arqueológico (en términos de conservación de cráneos completos), o a que no haya una gran cantidad de sitios representados en las muestras de las otras regiones (a excepción de Colombia, representada por dos sitios). Estudios futuros deberán mejorar el diseño muestral e incluir, por ejemplo, para Brasil, individuos de varios sitios arqueológicos de distintas regiones para poner a prueba la hipótesis de un modelo mixto (e.g., divergencia temprana y migraciones trasandinas) en pos de explicar la diversificación de los humanos en Sudamérica durante el Holoceno temprano y medio.