Introducción
Los datos etnohistóricos de comienzos del siglo XVI disponibles para la región del humedal del Paraná inferior (Díaz de Guzmán, 1836; Schmidl, 1903; Lopes de Souza, 1969, entre otros) señalan la presencia de distintos grupos étnicos tales como chanás, timbúes, querandíes, mbeguás y guaraníes, quienes además de mantener relaciones sociales entre sí y compartir aspectos organizativos sociales, económicos y simbólicos también interactuaban con grupos de áreas vecinas, como las regiones de Tandilia, Patagonia, la provincia de Córdoba, o bien de las costas de la actual república del Uruguay (Torres, 1911; Lothrop, 1932; Ceruti, 2000; Ceruti y González, 2007; Loponte, 2008; Mazza y Loponte, 2012; Acosta y Loponte, 2013; Fabra, 2013; Bonomo, Angrizani, Apollinaire y Noelli, 2015; Bonomo, Cabanillas y Montero, 2017; Bonomo et al., 2019).
A partir de 1527, con la instalación del fuerte Sancti Spiritu a la vera del río Carcarañá, en territorio de la actual provincia de Santa Fe, se inicia la dominación colonial en la región mesopotámica (Cocco et al., 2016). Este proceso produjo, entre otras consecuencias, la drástica disminución del número de nativos, la extinción de etnias, así como también la alteración del uso del espacio, de las redes de alianza y comercialización (González Lebrero, 2002; Tapia, 2002). No obstante, durante este periodo las poblaciones indígenas no fueron simples observadores de la dominación colonial hispánica; también habrían elaborado diversas estrategias de resistencia al adoctrinamiento religioso, al trabajo forzado y a la reducción en “pueblos de indios” (Lightfoot, 2005; Fradkin y Garavaglia, 2009). Ante la huida, muerte o relocalización de los nativos, las autoridades coloniales intentaron mantener un número aceptable de individuos que pudiesen sostener su continuidad, tanto laboral como económica. Por ello, mediante la persuasión o la fuerza redujeron indígenas que en muchas ocasiones provenían de lugares lejanos (Lightfoot, 2005; Tapia, Néspolo y Noya, 2015). Este aspecto resulta significativo a los fines del estudio que se expone en este trabajo, por cuanto los nuevos aportes poblacionales incidieron en la perduración de las identidades culturales previas, al producir procesos de etnogénesis en las nuevas generaciones.
Santiago del Baradero es una de las reducciones franciscanas instaladas en el Paraná medio e inferior que se encuentra mejor caracterizada a partir de la evidencia arqueológica del sitio Cementerio Indígena (CIB) y la disponibilidad de documentación escrita. Fue fundada por Hernando Arias de Saavedra en 1615, sobre las riberas del río Baradero y en el espacio que actualmente ocupa el centro cívico de la ciudad homónima, ubicada en la provincia de Buenos Aires. Este establecimiento misional originalmente estuvo integrado por individuos de grupos litorales como los chaná, mbeguá y guaraní y poco después de su fundación se sumaron algunos indígenas provenientes de otras regiones del país (González Lebrero, 2002; Tapia, 2002, 2014; Tapia, Néspolo y Noya, 2015; Cardozo, Tapia y Dejean, 2018).
La información documental disponible para CIB se correlaciona con evidencias arqueológicas. En efecto, se recuperaron tanto materiales indígenas como de procedencia europea formando parte del ajuar funerario de 14 individuos exhumados (Debenedetti, 1911). Entre los materiales indígenas, además de objetos típicos de algunos grupos prehispánicos del humedal del Paraná inferior (HPI) (e.g., cuentas de valvas de moluscos de río), también se distingue la presencia de discos de latón que presentan un diseño similar a los confeccionados por otros grupos poshispánicos del centro-oeste cordillerano, como los pehuenches del sur mendocino y norte neuquino (Tapia, Landa, De Rosa y Montanari, 2009; Tapia, 2014). Dichos hallazgos ponen en evidencia las estrechas relaciones de intercambio y las alianzas que mantenían entre sí los diferentes grupos indígenas durante los periodos poscoloniales, como así también la necesidad que las autoridades coloniales tuvieron de relocalizar individuos para mantener el número de población indígena y utilizar su fuerza de trabajo en pos del funcionamiento de los pueblos de indios y las encomiendas coloniales (González Lebrero, 2002; Fradkin y Garavaglia, 2009).
Acerca de los marcadores genéticos en arqueología
Para los estudios de genética de poblaciones el marcador más utilizado es el ADN mitocondrial (indicador de ancestría materna). Hasta el momento, mediante el análisis de las mutaciones puntuales presentes en este genoma ha sido posible distinguir 16 linajes o haplogrupos (Hg) mitocondriales amerindios fundadores. De ellos, algunos se encuentran distribuidos en todo el continente: A2, B2, C1b, C1c, C1d, C1d1, C4c, D1 y D4h3a; mientras que los restantes se encuentran restringidos a Norteamérica: A2a, A2b, D2a, D3, D4e1, X2a y X2g (Perego et al., 2010; de Saint Pierre et al., 2012a; Kashani et al., 2012). Los efectos de la deriva génica y otras fuerzas evolutivas que actuaron durante el poblamiento del continente estructuraron la distribución genética en diferentes proporciones a medida que se iban poblando nuevos territorios. De esta manera, se generaron linajes mitocondriales característicos (subhaplogrupos) derivados de los linajes continentales fundadores, cuya distribución se asocia a determinadas regiones (Perego et al., 2009; Perego et al., 2010; Bodner et al., 2012; de Saint Pierre et al., 2012a, 2012b; Gómez-Carballa et al., 2018).
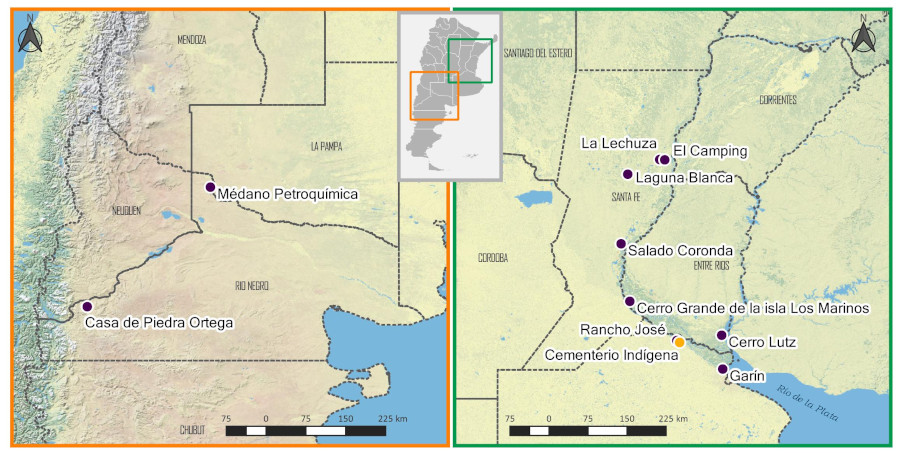
Los datos genéticos también han demostrado las consecuencias negativas de la conquista hispánica, dado que indican que hace aproximadamente 500 años, el tamaño efectivo de la población femenina se habría reducido en aproximadamente un 50% (O’Fallon y Fehren-Schmitz, 2011; Llamas et al., 2016). Por ello, para comprender los procesos demográficos generados por el impacto de la conquista europea en el HPI, resulta de interés explorar la variabilidad genética de los individuos asentados durante épocas prehispánicas como durante los primeros momentos de la dominación europea, en conjunto con los datos recabados desde la arqueología histórica y la etnohistoria. Para lograr este objetivo se busca integrar la información proveniente de los registros documentales de la época colonial con los datos genéticos actualmente disponibles para la reducción Santiago del Baradero (Cardozo et al., 2018) y aquellos provenientes de otros sitios del Paraná medio e inferior (PMI). Al considerar la presencia de los discos de latón con diseño de algunas poblaciones de la región centro-oeste del país, se analizan restos de individuos encontrados en esta última con el fin de comparar las frecuencias de Hg en ambas regiones (Figura 1). A la vez, se comparan los resultados genéticos obtenidos con los de ADN mitocondrial relevados de la bibliografía para otras regiones de interés.
Figura 1
Ubicación de los sitios arqueológicos de los cuales provienen las muestras utilizadas en el análisis comparativo de ADNa. En el recuadro de la derecha se indican: el sitio Cementerio Indígena de Baradero y los que corresponden a los sectores del río Paraná medio e inferior. En el recuadro de la izquierda se indican los sitios que corresponden al centro-oeste.
Materiales y métodos
Fuentes documentales
Se analizaron 18 fuentes escritas del Archivo General de Indias (AGI) que corresponden a la Audiencia de Charcas (ACH) y Audiencia de Buenos Aires (AB) e incluyen informes y cartas elevadas por diferentes funcionarios del gobierno de Buenos Aires a la Corona española y viceversa. Actualmente, dichas fuentes se encuentran disponibles en el repositorio documental del Museo Etnográfico, Facultad de Filosofía y Letras de la Universidad de Buenos Aires (AGI-ACH 1599, 1609, 1616, 1617, 1619, 1620, 1621-1622, 1622, 1626, 1628, 1664, 1673, 1678, 1690, 1708, 1718; AGI-AB 1679, 1691). Desde el punto de vista metodológico, las fuentes se clasificaron en dos grupos: cartas y expedientes (informes de funcionarios o autoridades eclesiásticas, disposiciones reales, pleitos sumarios, probanzas de méritos y servicios), lo cual permitió considerar los datos heterogéneos del repositorio documental (Tabla 1). Su análisis ha proporcionado información sobre el funcionamiento de la reducción Santiago del Baradero, desde su creación hasta su disolución administrativa como pueblo de indios en 1746 (Tapia et al., 2015). Especialmente para la problemática de estudio, resultan de interés las referencias sobre la forma de vida reduccional de los nativos, las características de las relaciones interétnicas y los movimientos de la población indígena.
Tabla 1
Número de fuentes del AGI consultadas y su distribución por categoría y cronología (las citas de las fuentes se listan al final del texto; Subt.: Subtotal; Exp.: Expedientes).
|
Archivo General de Indias (AGI) |
Número de fuentes consultadas |
Años que se referencian |
|
Subt. |
Exp. |
Cartas |
Siglo XVII |
Siglo XVIII |
|
1600-1650 |
1651-1700 |
1701-1750 |
|
Audiencia de Charcas (ACH) |
16 |
8 |
8 |
10 |
5 |
1 |
|
Audiencia de Buenos Aires (AU) |
2 |
1 |
1 |
- |
1 |
1 |
|
Totales |
18 |
9 |
9 |
10 |
6 |
2 |
Muestras arqueológicas
Dada la intención de comparar marcadores mitocondriales entre individuos nativos prehispánicos y poshispánicos que habitaron las costas del río Paraná cercanas a Baradero y del centro-oeste de la Argentina, se utilizaron y analizaron muestras de dichas regiones y agrupadas en dos grupos poblacionales: 29 muestras se incluyeron para la región del PMI y 6 muestras para la región centro-oeste (CO). Adicionalmente, los nuevos datos genéticos obtenidos se compararon con los de las 20 muestras previamente analizadas del CIB (Cardozo et al., 2018) (Figura 1, Tabla 2).
Todos los restos humanos analizados corresponden a colecciones museográficas y a registros arqueológicos recuperados por arqueólogos que actualmente desarrollan sus investigaciones en el país y mantienen sus colecciones en tránsito en sus lugares de trabajo.1 Para su estudio se ha cumplido con las normativas de la Ley 25.517/2001 y el Código de Ética del ICOM (suscripto por la República Argentina), respecto de las responsabilidades de los profesionales sobre los restos humanos. Asimismo, para realizar el estudio genético se han respetado las normativas establecidas en el Código de Ética Profesional promulgado en 2009 por la Asociación de Arqueólogos Profesionales de la República Argentina (AAPRA) y el Código Deontológico establecido por la Asociación de Antropología Biológica Argentina (Aranda, Barrientos y Del Papa, 2014). Es importante destacar que no existen comunidades originarias que se vinculen directamente con los individuos prehispánicos y poshispánicos tempranos recuperados.
Tabla 2
Número de muestras analizadas por sitio arqueológico y asignaciones temporales.
Extracción de ADN y determinación de haplogrupos mitocondriales
Las extracciones de ADN se realizaron en el Centro de Estudios Biomédicos, Biotecnológicos, Ambientales y de Diagnósticos (CEBBAD) de la Universidad Maimónides, bajo protocolos de control de la contaminación en un laboratorio específicamente dedicado a estos estudios. Se utilizaron tres protocolos a partir de polvo de huesos o dentina: 1- extracción orgánica con fenol:cloroformo:isoamílico (25:24:1) y purificación utilizando el equipo Bioneer®; 2- kit Gene Clean® for ancient DNA (MPBiomedicals); y 3- kit Investigator® (Quiagen). De cada método se realizaron al menos dos extracciones a partir de cada muestra.
La determinación de los Hg se realizó mediante polimorfismo de longitud de fragmentos de restricción (RFLP). Se utilizaron cebadores extraídos de la bibliografía: 1- para los Hg A, C y D los propuestos por Stone y Stoneking (1998) o por Fehren-Schmitz, Reindel, Cacigao, Hummel y Hermann (2010); 2- para el Hg B se recurrió a Stone y Stoneking (1998) o uno diseñado por nuestro equipo de investigación (Cardozo et al., 2018). Para la determinación de los Hg A, C y D se utilizaron las enzimas de restricción Hae III, Hinc II y Alu I respectivamente; mientras que en el Hg B se buscó la deleción de 9 pares de bases características. La reacción en cadena de la polimerasa y el tratamiento de los fragmentos con sus respectivas enzimas de restricción se llevaron a cabo como se describe en Cardozo et al. (2018).
Análisis estadístico comparativo
A partir de la información recabada de los registros arqueológicos, históricos y etnohistóricos se realizó un relevamiento bibliográfico de datos genéticos mitocondriales de poblaciones amerindias arqueológicas y contemporáneas (en estas últimas, se priorizaron comunidades nativas o pueblos pequeños organizados desde su fundación como pueblos de indios) afines a la región. La búsqueda se centró en las regiones pampeana, Pampa-Patagonia, serrana, Gran Chaco, Paraguay, Uruguay y sur de Brasil, y con esta información se confeccionó una base de datos con los correspondientes Hg mitocondriales. A partir de estos últimos se recalcularon las frecuencias, agrupando aquellas muestras que incluían poblaciones cercanas y relacionadas genéticamente, y eliminando muestras “indeterminadas” o correspondientes a linajes europeos (Tabla 3). Con los datos de frecuencias de Hg propios y de la bibliografía se realizó un análisis de coordenadas principales (ACP) con el programa Infostat (Di Rienzo et al., 2008) utilizando la distancia de Bray-Curtis con el fin de analizar las distancias genéticas entre las poblaciones.
Por otro lado, se evaluó la diferenciación genética entre las poblaciones con un análisis de la varianza molecular (AMOVA) utilizando 1000 permutaciones para el cálculo de los estadísticos F. Se realizó además una comparación de a pares entre poblaciones mediante estadísticos FST utilizando 1000 permutaciones con un nivel de significancia de 0,05. Para ambos análisis se utilizó el programa Arlequin v. 3.5 (Excoffier y Lischer, 2010).
Tabla 3
Frecuencia de los haplogrupos amerindios obtenidos de la búsqueda bibliográfica.
|
Temporalidad |
Conjunto muestral |
Región |
Siglas |
N |
A% |
B% |
C% |
D% |
Publicación |
|
Arqueológicas |
Arroyo Seco |
Sur de Buenos Aires |
AS |
10 |
10 |
30 |
40 |
20 |
Llamas et al. (2016) |
|
Figueiro (2013b) |
|
Uruguay |
Sureste y suroeste |
Urug |
23 |
17,4 |
39,1 |
34,8 |
8,7 |
Figueiro (2013a) |
|
Córdoba Antiguo |
Córdoba |
CbaA |
15 |
27 |
33 |
33 |
7 |
Nores y Demarchi (2011) |
|
Córdoba Intermedio |
CbaI |
18 |
17 |
22 |
39 |
22 |
Nores y Demarchi (2011) |
|
Norpatagonia andino |
Neuquén y Río Negro |
NPA |
4 |
25 |
- |
50 |
25 |
Crespo, Russo, Hajduk, Lanata y Dejean (2017a) |
|
Norpatagonia costa |
Río Negro |
NPC |
11 |
9 |
- |
9 |
82 |
Crespo, Favier Dubois, Russo Lanata y Dejean (2017b) |
|
Modernas |
Mataco Chaco |
Chaco |
MCh |
28 |
10,7 |
35,7 |
0 |
53,6 |
Torroni et al. (1993) |
|
Toba Chaco 1 |
TbCh |
71 |
14,1 |
50,7 |
4,2 |
31 |
Demarchi, Panzetta-Dutari, Motran, López de Basualdo y Marcellino (2001) |
|
Toba Chaco 2 |
Cabana, Merriwether, Hunley y Demarchi (2006) |
|
Toba Formosa |
Formosa |
TF |
26 |
27 |
34,6 |
3,8 |
34,6 |
Demarchi et al. (2001) |
|
Pilaga 1 |
MP |
122 |
8,2 |
41,8 |
25,4 |
24,6 |
Demarchi et al. (2001) |
|
Mataco Formosa |
|
Pilaga 2 |
Cabana et al. (2006) |
|
Toba Formosa 2 |
TW |
398 |
28 |
19,8 |
2,5 |
49,7 |
|
Wichi Chaco |
Chaco |
|
Wichi Chaco 2 |
Servini et al. (2013) |
|
Wichi Formosa |
Formosa |
WF |
67 |
8,9 |
56,7 |
19,4 |
15 |
Cabana et al. (2006) |
|
Mbyá-Guaraní |
Misiones |
MG |
45 |
48,9 |
33,3 |
8,9 |
8,9 |
Altuna y Demarchi (2003) |
|
Guaraní |
Misiones |
G |
145 |
42,1 |
14,5 |
4,8 |
38,6 |
Sala et al. (2010) |
|
Guaraní M’byá |
Paraná, Brasil |
Marrero et al. (2007) |
|
Guaraní Ñandeva |
Mato Grosso, Brasil |
GÑ |
56 |
82,1 |
0 |
16,1 |
1,8 |
Marrero et al. (2007) |
|
Mapuche |
Río Negro |
Mpch |
180 |
12,8 |
38,9 |
21,1 |
27,2 |
de Saint Pierre et al. (2012b) |
|
Mapuche 2 |
Goicoechea et al. (2001), Ginther et al. (1993) |
|
Tehuelche |
Chubut |
Tlch |
29 |
0 |
20,7 |
20,7 |
58,6 |
de Saint Pierre et al. (2012b) |
|
Villa Atamisqui |
Santiago del Estero |
VA |
53 |
17 |
11,3 |
51 |
20,7 |
Pauro, García, Nores y Demarchi (2013) |
|
Sumampa |
S |
28 |
21,5 |
7,1 |
32,1 |
39,3 |
Pauro et al. (2013) |
Resultados obtenidos
Análisis de las fuentes documentales: el contexto sociohistórico colonial
Cuando Hernandarias fundó la misión Santiago del Baradero en 1615 había logrado reunir en ella un número considerable de individuos de las etnias guaraní y chaná-mbeguás: “llegué a otra Reducción por nombre Santiago […] y hay en esta Reducción 250 indios” (AGI-ACH, 1616, f. 2). No obstante, en pocos años el número de nativos asentados fue disminuyendo y, según se infiere a partir de las fuentes escritas, es posible esbozar algunas de las posibles causas. Entre estas se destaca la alta mortalidad, producto de las epidemias y las continuas huidas por temor a contagiarse de las enfermedades. Al respecto, se indica: “habíase muerto de peste más de 60 y otros se habían ausentado del temor de ella” (AGI-ACH, 1621-1622, f. 14). Puede interpretarse cierta disconformidad con el modo de vida reduccional: “dichos naturales, unos más que otros, son pobrísimos […] y que los que son labradores tienen el sustento muy tasado y los demás viven como de milagro sustentándose con caza y pesquería” (AGI-ACH, 1619, f. 14). Por otra parte, se destaca la resistencia de los nativos a adoptar la sedentarización: “no tienen casa ni asiento” (AGI-ACH, 1599, f. 3). La disminución de individuos en las reducciones también puede ser causa de los malos tratos que los encomenderos daban a los nativos reducidos: “haber hecho mucha cantidad de yeguas por mandado de Bartolomé Pintos su administrador y no haberles pagado nada por ello y haber sido tratado mal del dicho administrador” (AGI-ACH, 1620, f. 10).
Debido a la paulatina pérdida de mano de obra indígena se establecieron distintas medidas de control. Entre ellas se destaca la prohibición de que las mujeres de un pueblo de indios se unieran en matrimonio con nativos de otros establecimientos coloniales, ya que se deberían mudar al lugar donde residía el esposo. Si bien el rey Felipe III reclamó que no se prohibieran dichas uniones, estas continuaron de manera solapada:
He sido informado que a las indias de esa provincia de los repartimientos que estan encomendados no se les deja libertad para casarse porque si se casa la india de un repartimiento con indio de otro el marido lleva a la muger y el encomendero porque no le saque la india de su encomienda impide estos matrimonios a lo qual no es justo que se de lugar. (AGI-ACH, 1609, f. 1)
Además, se prohibieron las salidas de los nativos: “no permitiendo se saquen de él ninguno, ni menos indias, muchachos ni muchachas con ningún pretexto” (AGI-ACH, 1708, f. 9). Con esta decisión - que afectaba particularmente a las mujeres dado que ellas engendrarían la futura fuerza de trabajo- se intentaba preservar la mano de obra joven. En 1688 se realizó un padrón de indios en la reducción Santiago del Baradero; en él se consigna que todas las mujeres habían nacido en la reducción o en la región (AGI-ACH, 1690). Este dato es relevante al evaluar la diversidad genética mitocondrial, dada la transmisión por línea materna de este ADN. También se identificaron individuos provenientes de otras regiones del país, cuyos aportes poblacionales fueron registrados en un padrón confeccionado entre 1684 y 1686:
mando se haga padrón de todas las dichas familias de indios con distinción de sus naturalezas, sexos y edades poniendo con separación los que han sido descendientes de la misma reducción y los que son agregados expresando las familias de los unos y los otros. (AGI-ACH, 1690, f. 4)
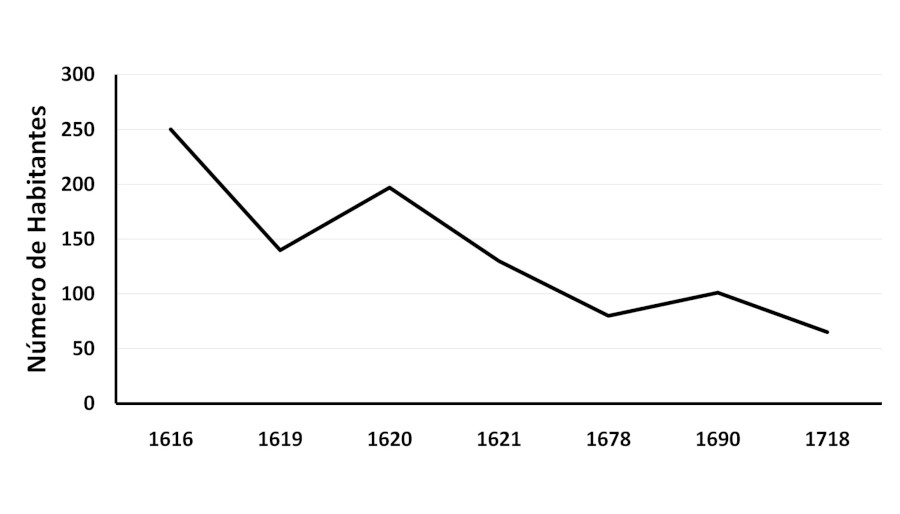
Del total de 101 indígenas empadronados, todas las mujeres eran nacidas en la reducción o de áreas cercanas, mientras que nueve de los varones adultos eran foráneos y estaban casados con mujeres que habían nacido en la reducción: cinco provenían de regiones alejadas como Santiago del Estero (2), Cuzco (1) o Paraguay (2); mientras que los otros de Santa Fe (2), Corrientes (1) o simplemente asentado como Guaraní (1) (AGI-ACH, 1690). En la Figura 2 se muestra la fluctuación y la tendencia decreciente del número de indígenas asentados en Santiago del Baradero a lo largo de 102 años.
Figura 2
Variación en el número de individuos en Santiago del Baradero según las fuentes documentales. AGI-ACH: 1616; 1619; 1620; 1621-1622; 1678; 1690; 1718.
Datos genéticos: rendimiento de las extracciones
De los 45 restos analizados se realizaron 228 extracciones aplicando las tres técnicas detalladas. Se analizaron tres factores: método de extracción, tipo de resto y región geográfica. Los mejores resultados se obtuvieron con el protocolo de fenol:cloroformo:isoamílico con piezas dentales y con las muestras de la región centro-oeste respectivamente (Tabla 4).
Tabla 4
Rendimiento de las extracciones realizadas en muestras de diente y hueso de todos los sitios analizados con los distintos métodos.
|
Conjunto muestral |
Tipo de pieza |
Gene Clean® |
Extracción orgánica |
Investigator® |
|
Total |
Efectividad |
Total |
Efectividad |
Total |
Efectividad |
|
Cementerio indígena |
Dental |
10 |
40% |
19 |
36,80% |
14 |
57,10% |
|
Ósea |
20 |
30% |
18 |
50% |
15 |
26,70% |
|
Paraná medio e inferior |
Dental |
28 |
21,40% |
43 |
28% |
15 |
33,30% |
|
Ósea |
9 |
0% |
19 |
26,30% |
2 |
0% |
|
Centro oeste |
Dental |
8 |
25% |
6 |
83,33% |
2 |
100% |
Resultados generales de RFLP
Además de las muestras ya publicadas del sitio CIB, se pudo determinar el Hg de dos nuevas muestras (CIB2 y CIB4), mientras que para PMI y CO fue posible determinar el Hg para cinco y tres muestras respectivamente. En total, junto con los datos de Cardozo et al. (2018), se cuenta con los Hg de 19 muestras, de las cuales 18 son amerindias y la restante no pudo incluirse dentro de los linajes nativos (Figura 3, Tabla 5). Es de destacar que esta última es poshispánica (CIB5).
Figura 3
Datos de haplogrupos obtenidos a partir de muestras de distintos sitios: A) Rendimiento en los distintos conjuntos; y B) Hg encontrados en cada conjunto. CIB: Cementerio Indégena de Baradero, PMI: Paraná medio e inferior.
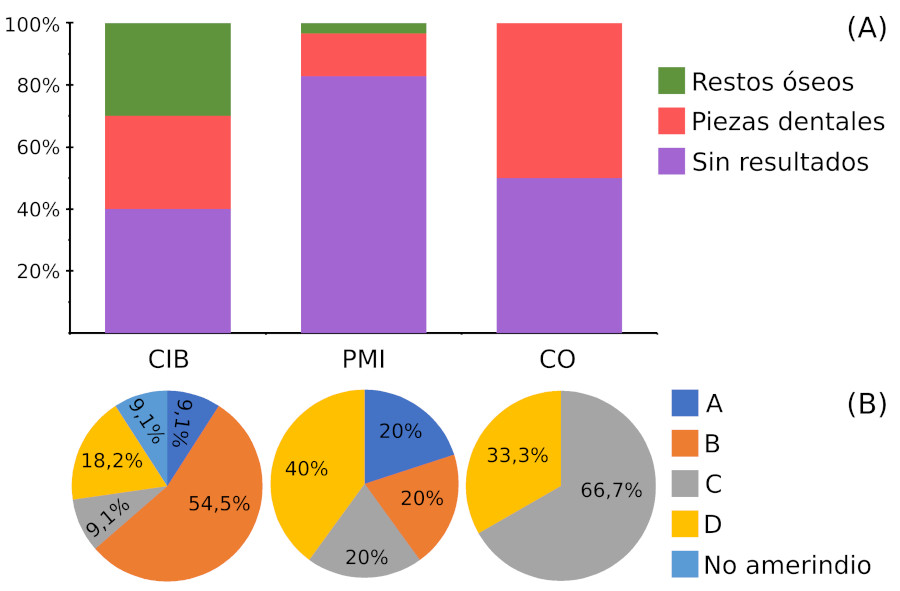
Tabla 5
Hg obtenidos para los distintos sitios analizados
|
Área de procedencia |
Sitio |
Muestra |
Código original de inventario |
Hg |
Publicación |
|
Baradero |
Cementerio Indígena de Baradero |
CIB2 |
CIB7 |
D |
Esta publicación |
|
CIB3 |
CIB120 |
No amerindio |
Cardozo et al. (2018) |
|
CIB8 |
CIB79 |
A |
|
CIB10 |
CIB121 |
C |
|
CIB11 |
CIB123 |
B |
|
CIB14 |
CIB298 |
B |
Esta publicación |
|
CIB16 |
CIB155 |
B |
Cardozo et al. (2018) |
|
CIB17 |
CIB185 |
B |
|
CIB18 |
CIB186 |
B |
|
CIB20 |
CIB347 |
D |
|
CIB21 |
CIB352 |
B |
|
Paraná Medio e Inferior |
Los Marinos |
LM5bis |
2/39-5 bis |
C |
Esta publicación |
|
Cerro Lutz |
CL9 |
CL9 |
B |
|
La Lechuza |
SC-A1 |
A1 |
D |
|
El Camping |
SC-C1 |
C1 |
D |
|
Río Salado-Coronda II |
RF2 |
RSC II R16 |
A |
|
Centro-Oeste |
Médano Petroquímica |
MP-2 |
Indiv. 6 H4 |
C |
|
MP-5 |
Sin Nº H5 Mandíbula 4 |
D |
|
Casa de Piedra Ortega |
CE5 |
CPO |
C |
En las muestras de CIB se encontró mayor presencia del Hg B2, mientras que en PMI dos pertenecen a D, y las restantes, a los otros linajes amerindios. Finalmente, los individuos agrupados en CO presentan solo Hg C y D (Figura 3). A partir de estos resultados, en la Tabla 6 se indican las frecuencias de los linajes amerindios mitocondriales obtenidos.
Tabla 6
Frecuencia de haplogrupos amerindios obtenidos a partir de las muestras analizadas.
|
Conjunto muestral |
Región |
Siglas |
n |
A% |
B% |
C% |
D% |
|
Cementerio Indígena de Baradero |
Baradero, Buenos Aires |
CIB |
10 |
10 |
60 |
10 |
20 |
|
Río Paraná medio e inferior |
Buenos Aires, Santa Fe y Entre Ríos |
PMI |
5 |
20 |
20 |
20 |
40 |
|
Región Centro-Oeste |
La Pampa y Neuquén |
CO |
3 |
0 |
0 |
66,7 |
33,3 |
Comparación de datos genéticos en escala interregional
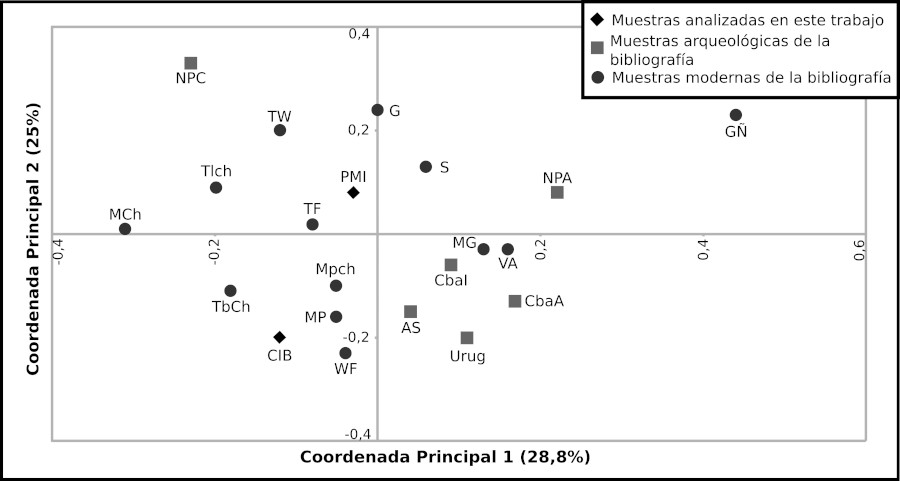
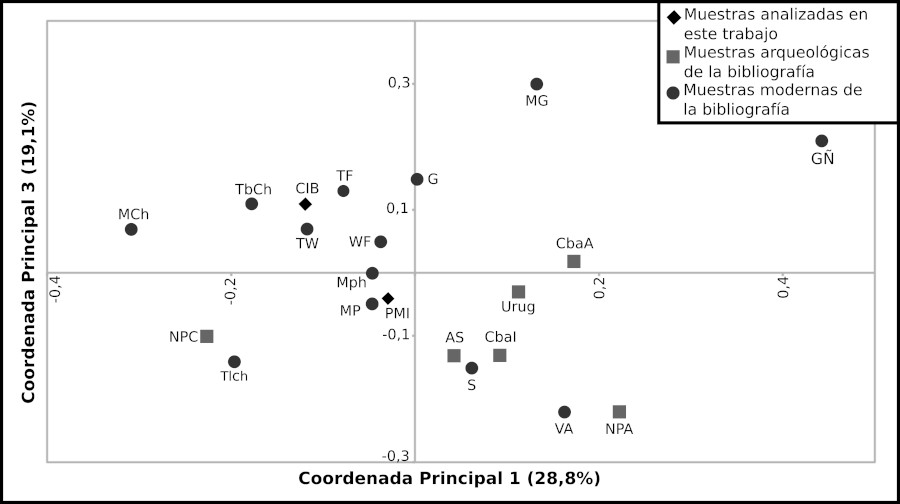
En el ACP se observó que los tres primeros ejes explican el 72,9% de la variabilidad existente entre los conjuntos analizados. Los coeficientes de correlación indican que la coordenada principal 1 discrimina en función de los Hg A y D; la coordenada principal 2 con el Hg B; y la coordenada principal 3, con el Hg C (Tabla 7). En la Figura 4 se observa que las muestras de CIB se encuentran más cercanas a las poblaciones wichi de Formosa (WF), tobas de Chaco (TbCh) y las poblaciones pilaga y matacos de Formosa (MP). Por su parte, las muestras prehispánicas del PMI se agrupan cercanas a las muestras tobas de Formosa (TF). Finalmente, dado el bajo número de resultados de Hg para CO, estos no se incluyeron en ninguno de los análisis estadísticos.
Tabla 7
Coeficientes de correlación entre los Hg y las tres primeras coordenadas principales (*: p-valor ≤0,001)
|
Hg |
Coordenada principal 1 |
Coordenada principal 2 |
Coordenada principal 3 |
|
A |
0,70* |
0,42 |
0,57 |
|
B |
-0,39 |
-0,83* |
0,38 |
|
C |
0,57 |
-0,28 |
-0,76* |
|
D |
-0,75* |
0,60 |
-0,26 |
Figura 4
Distribución de los conjuntos en función de las coordenadas principales 1 y 2. Cementerio Indígena de Baradero (CIB); Río Paraná medio e inferior (PMI); Arroyo Seco (AS); Uruguay (Urug); Córdoba Antiguo (CbaA); Córdoba Intermedio (CbaI); Norpatagonia Andino (NPA); Norpatagonia Costa (NPC); Mataco Chaco (MCh); Toba Chaco 1 y 2 (TbCh); Toba Formosa (TF); Mataco Formosa, Pilaga 1 y 2 (MP); Toba Formosa 2, Wichi Chaco 1 y 2 (TW); Wichi Formosa (WF); Mbya-Guaraní (MG); Guaraní y Guaraní M´Byá (G); Guaraní Ñandeva (GÑ), Mapuche 1 y Mapuche 2 (Mpch), Tehuelche (Tlch); Villa Atamisqui (VA); Sumampa (S).
En la Figura 5, la muestra de CIB se ubica cercana al conjunto de tobas de Chaco (TbCh), tobas de Formosa (TF) y la población toba de Formosa y wichi de Chaco (TW). Por su parte, los individuos de la región PMI se ubican cercanos a la población pilaga y matacos de Formosa (MP) y las poblaciones mapuche de Río Negro.
Figura 5
Distribución de los conjuntos en función de las coordenadas principales uno y tres. Cementerio Indígena de Baradero (CIB); Río Paraná Medio e Inferior (PMI); Arroyo Seco (AS); Uruguay (Urug); Córdoba Antiguo (CbaA); Córdoba Intermedio (CbaI); Norpatagonia Andino (NPA); Norpatagonia Costa (NPC); Mataco Chaco (MCh); Toba Chaco 1 y 2 (TbCh); Toba Formosa (TF); Mataco Formosa, Pilaga 1 y 2 (MP); Toba Formosa 2, Wichi Chaco 1 y 2 (TW); Wichi Formosa (WF); Mbya-Guaraní (MG); Guaraní y Guaraní M´Byá (G); Guaraní Ñandeva (GÑ), Mapuche 1 y Mapuche 2 (Mpch), Tehuelche (Tlch); Villa Atamisqui (VA); Sumampa (S)
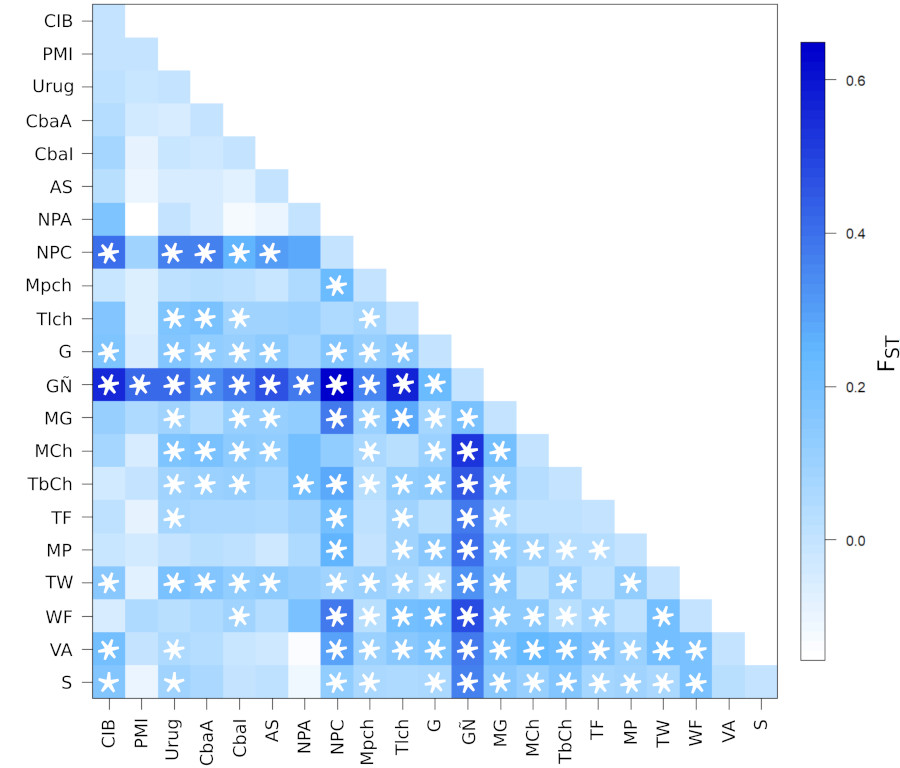
En el AMOVA se observó un porcentaje estadísticamente significativo de variación del 11,45% entre las distintas poblaciones (p ≤0,001). En concordancia con el ACP, en la Matriz de Fst y de p-valores se observa que CIB se encuentra más distanciado de poblaciones antiguas de Norpatagonia Costa (NPC), Guaraníes (G y GÑ), Tobas y Wichi (TW) y de Santiago del Estero (VA y S). Por su parte, PMI solo se distancia significativamente de los Guaraní Ñandeva (GÑ) (Figura 6).
Es interesante destacar que, si bien el análisis de coordenadas principales muestra distanciamiento genético entre las poblaciones prehispánicas PMI y poshispánica CIB, la comparación de a pares indica que tal diferenciación no es significativa.
Figura 6
Comparación de a pares (Fst) de las poblaciones analizadas en este trabajo y aquellas obtenidas de la búsqueda bibliográfica (* equivale a un p<0,05). Cementerio Indígena de Baradero (CIB); Río Paraná Medio e Inferior (PMI); Arroyo Seco (AS); Uruguay (Urug); Córdoba Antiguo (CbaA); Córdoba Intermedio (CbaI); Norpatagonia Andino (NPA); Norpatagonia Costa (NPC); Mataco Chaco (MCh); Toba Chaco 1 y 2 (TbCh); Toba Formosa (TF); Mataco Formosa, Pilaga 1 y 2 (MP); Toba Formosa 2, Wichi Chaco 1 y 2 (TW); Wichi Formosa (WF); Mbya-Guaraní (MG); Guaraní y Guaraní M´Byá (G); Guaraní Ñandeva (GÑ), Mapuche 1 y Mapuche 2 (Mpch), Tehuelche (Tlch); Villa Atamisqui (VA); Sumampa (S).
Discusión y conclusiones
La recuperación general del 40% de ADNa resulta relevante al considerar los procesos diagenéticos que actúan sobre restos rescatados de regiones húmedas, los cuales dificultan la preservación y, por ende, recuperación de ADNa (Hofreiter et al., 2015). Estos resultados se obtuvieron mayoritariamente con el protocolo de extracción orgánica y con las piezas dentales. En el primer caso, la extracción resultó favorecida por la posibilidad de utilizar mayor cantidad de muestra y por la flexibilidad para modificar variables metodológicas; mientras que en las piezas dentales la cubierta de esmalte ofrece una protección adicional al ADN frente a los procesos de descomposición del material orgánico.
Respecto de los datos históricos que proporcionan los documentos escritos, es importante recordar que fueron generados por funcionarios coloniales desde una perspectiva sesgada, la cual intentaba justificar el proceso de colonización y las relaciones de dominación aplicadas sobre los grupos nativos (Solodkow, 2009; Tapia, 2014). No obstante, acotando las fuentes escritas a su contexto histórico particular y desde un enfoque etnohistórico crítico, es posible considerar datos significativos para la cuestión del impacto que habría producido la conquista en la variabilidad genética de las poblaciones nativas del área de estudio.
En tal sentido, en la información documental se observan ciertas características demográficas propias de la reducción Santiago del Baradero que apoyarían lo observado en la Comparación de a Pares entre el grupo prehispánico de PMI y el poshispánico CIB. Por ejemplo, en el padrón de 1688 se especifica que, de la población total, solo nueve nativos varones eran foráneos procedentes del nordeste y noroeste del país; además, se hace referencia a los controles demográficos coloniales que habrían restringido la movilidad de las mujeres indígenas fuera de la misión. De esta manera, resulta plausible inferir que las estrategias de reducir diferentes grupos étnicos de la región dentro de un pueblo de indios no habrían producido un impacto genético de magnitud como podría llegar ser el surgimiento de un efecto fundador2 que podría haber generado la diferenciación entre las poblaciones pre- y poshispánicas. No obstante, el tamaño reducido de la muestra y el tipo de marcador mitocondrial utilizado podrían ocultar el accionar de otros mecanismos microevolutivos.
Si bien el grupo mapuche actual muestra cercanía con PMI (Figura 5) y en menor medida con CIB (Figura 4), estos conjuntos se encuentran alejados genéticamente de otras poblaciones patagónicas relevadas de la bibliografía, como Norpatagonia Costa (NPC), Norpatagonia Andino (NPA) y Tehuelches (Tlch), así como con la mayoría de los datos de restos arqueológicos provenientes de otras regiones de Patagonia, donde los Hg mayoritarios son C y D (Crespo et al., 2017a y b). En tal sentido, si bien las muestras de CO no se incluyeron en los análisis estadísticos, cabe destacar que los únicos Hg encontrados hasta el momento corresponden a C y D. No obstante, existe evidencia arqueológica del Hg B2 hallada en cinco individuos rescatados de distintos entierros de la cuenca del Lago Salitroso en la provincia de Santa Cruz (301-664 AP) (Arencibia et al., 2019). Adicionalmente, el análisis de poblaciones nativas contemporáneas de Norpatagonia permitió identificar la presencia del subhaplogrupo B2i2, cuyas frecuencias son elevadas en poblaciones mapuches de Chile y Argentina -26,3% y 38,9% respectivamente (de Saint Pierre et al., 2012a, b)-. Claramente, los datos obtenidos para CO no son determinantes de la ausencia del Hg B2 en esta región o de los posibles movimientos poblacionales, pero para indagar en estos tópicos sería necesario incrementar el número de secuencias disponibles para restos arqueológicos del CO, ya que no se corresponden con lo encontrado actualmente en el área o con las poblaciones arqueológicas que se encontraban más al sur de Patagonia.
En el ACP, ninguno de los conjuntos arqueológicos del PMI, CIB y de Uruguay se ubican cercanos a los grupos guaraníes (Altuna y Demarchi, 2003; Marrero et al., 2007; Sala et al., 2010). Esto podría deberse a la lejanía geográfica de estos últimos con respecto al HPI, sumado a ciertas características del grupo guaraní tales como su movilidad residencial, que generaba sucesivos efectos fundadores y contactos con otras etnias; y/o el cuello de botella sufrido luego de la conquista europea (Melo dos Santos, 2015). Así, el acervo génico de los grupos guaraníes modernos analizados puede no ser representativo de aquellos que arribaron al Río de La Plata hace 200 a 300 años previos a la llegada de los conquistadores europeos (Loponte, Acosta y Musali, 2004; Silva Noelli, 2004).
En uno de los individuos de CIB no se pudo determinar si su Hg es amerindio; sin embargo, en estudios posteriores de secuenciación se observaron mutaciones típicas del Hg C1 (datos aún inéditos). Esto ya ha sido observado en Torres et al. (2006), donde individuos de poblaciones nativas de Colombia, definidos como indeterminados por RFLP, portaban mutaciones propias del linaje C en la Región Hipervariable; y en Motti et al. (2009), donde encontraron la misma discordancia al momento de analizar muestras cosmopolitas de Mendoza y San Juan. Al respecto, se ha sugerido la ocurrencia de una mutación reversa3 del sitio de corte de Hinc II, lo que generó la pérdida de este.
Si bien existe un sesgo en los resultados genéticos obtenidos, dado el número de muestras y la metodología empleada, resulta de interés destacar la alta frecuencia de B (60%) en el acervo génico local. Dada la diversidad de subhaplogrupos B existentes en la región, B2i2 en Norpatagonia (de Saint Pierre et al., 2012a), B2y2a1a con dispersión por el Gran Chaco argentino o B2b3 en la ruta atlántica de Sudamérica (Gómez-Carballa et al., 2018), podrían ser varios los escenarios para los movimientos poblacionales que se dieron en el Paraná inferior según este Hg. No obstante, considerando la información arqueológica disponible respecto del flujo poblacional de grupos indígenas de la región central del país (Córdoba y Gran Chaco) hacia el PMI (Ceruti, 2000; Fabra, 2013), se podría inferir la existencia de relaciones de ancestralidad genética con las poblaciones modernas del Gran Chaco y Córdoba extraídas de la bibliografía.
Continuando con esta línea de análisis, dado que los censos presentes en las fuentes documentales no refieren la presencia de individuos provenientes de la región del centro de Argentina (porque la mayoría de los individuos ingresados en la reducción provendrían de grupos que habitaban la región del HPI), el acervo génico observado podría deberse, principalmente, a movimientos poblacionales previos a la llegada europea. Esto podrá corroborarse cuando se disponga de secuencias de muestras del PMI.
Por ello, la presencia de artefactos de latón en el ajuar funerario de los enterratorios del CIB -aunque puede responder a procesos de intercambio cultural interétnico-, desde la perspectiva genética continúa siendo una línea de investigación abierta a cuya resolución podremos acercarnos aumentando el número de muestras analizadas y logrando su secuenciación.
Si bien los datos arqueogenéticos obtenidos hasta el momento indican que en Santiago del Baradero las normativas coloniales no habrían producido un cambio significativo en el acervo génico del grupo indígena reducido, respecto de la población prehispánica preexistente, es necesario optimizar los métodos de extracción de ADNa en restos óseos y dentales afectados por las condiciones ambientales del HPI, para incrementar el número de muestras que permitan contrastar estos resultados y reforzar su confiabilidad. No obstante, estos primeros avances proporcionan información relevante para comprender algunos de los posibles efectos de la colonización hispánica en los pueblos originarios del pasado.
El avance en el estudio genético junto con el abordaje desde otras áreas de la bioarqueología y el cruzamiento con los datos de las fuentes documentales facilitará la postulación de hipótesis sobre el impacto antropobiológico de la conquista en las poblaciones nativas.
Agradecimientos
Agradecemos a la doctora M. Fernández, a los doctores O. Mendonça, M. Arrieta y a la doctora A. Aguerre, quienes gentilmente nos cedieron las muestras obtenidas en sus respectivas investigaciones. Asimismo, agradecemos los subsidios otorgados por la ANPCyT-FONCyT, el aporte del CONICET a través del otorgamiento de una beca doctoral al Dr. Darío Cardozo, el apoyo económico de la Fundación de Historia Natural Félix de Azara, Fundación Científica Felipe Fiorellino y por las secretarías de Ciencia y Técnica de la Universidad Nacional de Luján (Proyectos de investigación del Departamento de Ciencias Sociales) y de la Universidad de Buenos Aires (Proyectos UBACYT de la Facultad de Filosofía y Letras). Agradecemos especialmente a los revisores, cuyos aportes y correcciones enriquecieron de manera sustancial este trabajo.
Referencias bibliográficas
Acosta, A., Loponte, D. y Tchilinguirián, P. (2010). Análisis comparativo sobre la estructura y los procesos de formación de los depósitos arqueológicos en el humedal del río Paraná inferior (delta del Paraná). En G. Cocco y M. R. Feuillet Terzaghi (Eds.). Arqueología de cazadores recolectores en la Cuenca del Plata (pp. 191-208). Argentina: Centro de Estudios Hispanoamericanos.
Acosta, A. y Loponte, D. (2013). Complejidad social y estrategias de subsistencia de las poblaciones cazadoras-recolectoras del humedal del Paraná inferior. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano
(Series Especiales), 1(4), 60-74.
Acosta, A. y Mazza, B. (2016). Restos óseos humanos y faunísticos: su relación en el espacio mortuorio en contextos de cazadores-recolectores del humedal del Paraná Inferior (Argentina). Pesquisas Antropología, 72, 185-207.
Altuna, M. E. y Demarchi, D. A. (2003). Haplogrupos mitocondriales entre los mbya-guaraní de la provincia de Misiones. En Resúmenes Sextas Jornadas de Antropología Biológica, Revista Argentina de Antropología Biológica, 5(1), 115. Recuperado de https://revistas.unlp.edu.ar/raab/article/view/360/215
Ammann, M., Arrieta, M., Croatto, M. C., Bernardi, L., Mendonça, O. y Bordach, M. (2010). Descripción e interpretación de marcadores bioarquelógicos del sitio Médano Petroquímica, departamento de Puelén, provincia de La Pampa. En M. Berón, L. Luna, M. Bonomo, C. Montalvo, C. Aranda y M. Carrera Aizpitarte (Eds.). Mamül Mapu: Pasado y Presente desde la Arqueología Pampeana (pp. 105-112). Buenos Aires: Del Espinillo.
Aranda, C., Barrientos, G. y Del Papa, M. C. (2014). Código deontológico para el estudio, conservación y gestión de restos humanos de poblaciones del pasado. Revista Argentina de Antropología Biológica , 16(2). doi: 10.17139/raab.2014.00016.02.05
Arencibia, V., Crespo, C. M., García Guraieb, S., Russo, M. G., Dejean, C. B. y Goñi, R. (2019). Análisis genético poblacional de grupos cazadores recolectores del holoceno tardío del Lago Salitroso (Santa Cruz, Argentina). Revista Argentina de Antropología Biológica , 21(2). doi: 10.24215/18536387e004
Bodner, M., Perego, U. A., Huber, G., Fendt, L., Röck, A. W., Zimmermann, B., Olivieri, A., Gómez-Carballa, A., Lancioni, H., Angerhofer, N., Bobillo, M. C., Corach, D., Woodward, S. R., Salas, A., Achilli, A., Torroni, A., Bandelt, H. y Parson, W. (2012). Rapid coastal spread of first americans: Novel insights from South America’s southern cone mitochondrial genomes. Genome Research, 22(5), 811-820.
Bonomo, M., Angrizani, R., Apollinaire, E. y Noelli, F. (2015). A model for the Guaraní expansion in the La Plata Basin and littoral zone of southern Brazil. Quaternary International, 356, 54-73.
Bonomo, M., Cabanillas, E. y Montero, R. (2017). Archaeometallurgy in the Paraná Delta (Argentina): composition, manufacture, and indigenous routes. Journal of Anthropological Archaeology, 47, 1-11.
Bonomo, M., Di Prado, V., Silva, C., Scabuzzo, C., Ramos Van Raap, M., Castiñeira, C., Colobig, M. y Politis, G. (2019). Las poblaciones indígenas prehispánicas del río Paraná inferior y medio. Revista del Museo de La Plata, 4, 585-620.
Cabana, G., Merriwether, D., Hunley, K. y Demarchi, D. (2006). Is the genetic structure of Gran Chaco populations unique? Interregional perspective on native South American mitochondrial DNA variation. American Journal of Physical Anthropology, 131, 108-119.
Cardozo, D., Tapia, A. y Dejean, C. (2018). Datos genéticos e isotópicos de individuos vinculados a la reducción franciscana de Baradero (siglo XVII). Revista Arqueología Histórica Argentina y Latinoamericana, 12, 527-558.
Ceruti, C. (2000). Ríos y praderas: los pueblos del litoral. En M. Tarragó (Comp.). Nueva Historia Argentina, Los pueblos originarios y la conquista, Tomo 1 (pp. 105-146). Buenos Aires: Sudamericana.
Ceruti, C. y González, M. (2007). Modos de vida vinculados con ambientes acuáticos del nordeste y pampa bonaerense de argentina. Relaciones de la Sociedad Argentina de Antropología Biológica, 32, 101-140.
Cocco, G., Letieri, F., Frittegotto, G., Pasquali, C., Azkarate, A., Sanchez-Pinto, I., Escribano-Ruiz, S. y Benedet, V. (2016). Sancti Spiritus, 1527-1529. Aportes al estudio de los primeros asentamientos en Sudamérica. En L. M. Calvo y G. Cocco (Comps.). Primeros asentamientos españoles y portugueses en la América central y meridional, siglos XVI y XVII (pp. 201-2019). Santa Fe: Ediciones Universidad Nacional del Litoral.
Cornero, S. (2014). El Camping: Rescate y cronologías para un sitio Goya Malabrigo tardío, Alejandra, Dpto. San Javier, Provincia de Santa Fe. En C. López, L. Bulacio y M. Migliaro (Comps.). Ciencia y Tecnología: Divulgación de la Producción Científica y Tecnológica UNR (pp. 507-511). Rosario: Editorial de la Universidad Nacional de Rosario.
Cornero, S. (2016). Sitio La Lechuza: Aportes para la investigación en la arqueología norte de Santa Fe. En Resúmenes del XIX Congreso Nacional de Arqueología Argentina (pp. 480-484). Tucumán: Universidad Nacional de Tucumán.
Crespo, C., Russo, M., Hajduk, A., Lanata, J. y Dejean, C. (2017a). Variabilidad mitocondrial en muestras precolombinas de la Patagonia argentina. Hacia una visión de su poblamiento desde el ADN antiguo. Revista Argentina de Antropología Biológica , 19, 1-21.
Crespo, C., Favier Dubois, C., Russo, M., Lanata, J. y Dejean, C. (2017b). First analysis of ancient mtDNA genetic diversity in Northern coast of Argentinean Patagonia. Journal of Archaeological Science: Reports, 12, 91-98.
Curtis, H., Barnes, N., Schnek, A. y Massarini, A.(2015). Invitación a la biología en contexto social, 7ma. ed. Buenos Aires: Editorial Médica Panamericana
de Saint Pierre M., Gandini, F., Perego, U. A., Bodner M., Gómez-Carballa, A., Corach, D., Angerhofer, N., Woodward, S. R., Semino, O., Salas, A., Parson, W., Moraga, M., Achilli, A., Torroni, A. y Olivieri, A. (2012a). Arrival of paleo-indians to the southern cone of south America: new clues from mitogenomes. PLoS ONE, 7(12), e51311.
de Saint Pierre, M., Bravi, C., Motti, J., Fuku, N., Tanaka, M., Llop, E., Bonatto, S. y Moraga, M. (2012b). An Alternative Model for the Early Peopling of Southern South America Revealed by Analyses of Three Mitochondrial DNA Haplogroups. PLoS ONE , 7(9), e43486. doi:10.1371/journal.pone.0043486
Debenedetti, S. (1911). Noticia sobre un cementerio indígena de Baradero. Revista de la Universidad de Buenos Aires, Publicaciones de la Sección Antropológica, 13, 435-448.
del Rio, P., Cornero, S., Ceruti, C. N. y Echegoy, C. (2016). Arqueología de los Bajos Submeridionales: sitios con hornos de tierra cocida en la localidad arqueológica Laguna La Blanca (La Criolla, Departamento San Justo, provincia de Santa Fe). Revista de Antropología del Museo de Entre Ríos, 2(2), 67-82.
Demarchi, D., Panzetta-Dutari, G., Motran, C., López de Basualdo, M. y Marcellino, J. (2001). Mitochondrial DNA haplogroups in Amerindian populations from the Gran Chaco. American Journal of Physical Anthropology , 115, 199-203.
Díaz de Guzman, R. (1836 [1612]). Historia Argentina. Del descubrimiento, población y conquista de las provincias del Rio de la Plata. En P. de Angelis, Colección de obras y documentos relativos a la historia antigua y moderna relativos a las provincias del Rio de la Plata Tomo I. Buenos Aires: Imprenta del Estado.
Di Rienzo, J., Casanoves, F., Balzarini, M., González, L., Tablada, M. y Robledo, C. (2008). InfoStat Software Estadístico, versión 2008 Córdoba: Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba.
Excoffier, L. y Lischer, H. (2010). Arlequin suite ver. 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Molecular Ecology Resources, 10, 564-567.
Fabra, M. (2013). Variación epigenética craneofacial y dinámica evolutiva de poblaciones humanas del norte de la región pampeana durante el Holoceno tardío. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano
. Series Especiales, 1(1), 75-86.
Fehren-Schmitz, L., Reindel, M., Cacigao, E., Hummel, S. y Herrmann, B. (2010). Pre-columbian population dynamics in coastal Southern Peru: a diachronic investigation of mtDNA patterns in the Palpa region by ancient DNA analysis. American Journal of Physical Anthropology , 141, 208-221.
Fernández, M. (2001). La casa de piedra de Ortega (Pcia. de Río Negro) I. La estratigrafía. Relaciones de la Sociedad Argentina de Antropología, 26, 261-284.
Figueiro, G. (2013a). Continuidad temporal en la composición genética de las poblaciones indígenas del Uruguay. Colección Avances de Investigación, Tomo Docentes (pp. 141-159). Montevideo: Facultad de Humanidades y Ciencias de la Educación.
Figueiro, G. (2013b). Estudios de la Variación Temporal y Espacial en el ADN Mitocondrial de las Poblaciones Prehistóricas de la Región Pampeana (tesis de doctorado). Área Biología Universidad de la República, Montevideo, Uruguay.
Fradkin, R. y Garavaglia, J. (2009). La Argentina colonial: El Río de la Plata entre los siglos XVI y XIX Buenos Aires: Siglo XXI.
Galligani, P., Feuillet Terzaghi, M. y Barrientos, G. (2016). Los entierros humanos del sitio Río Salado-Coronda II: una aproximación tafonómica a los procesos de modificación ósea postdepositacional en el centro-este de la provincia de Santa Fe, República Argentina. Intersecciones en Antropología, 17(2), 187-200.
Gaspary, F. (1950). Investigaciones arqueológicas y antropológicas en un cerrito de la Isla los Marinos (Provincia de Entre Ríos). Revista del Instituto de Arqueología, Lingüística y Folklore “Dr. Pablo Cabrera“, 23, 3-66.
Ginther, C., Corach, D., Penacino, G. A., Rey, J. A., Carnese, F. R., Hutz, M. H., Anderson, A., Just, J., Salzano, F. M. y King, M. C. (1993). Genetic variation among the Mapuche Indians from the Patagonian region of Argentina: Mitochondrial DNA sequence variation and allele frequencies of several nuclear genes. En S. D. J. Pena, R. Chakraborty, J. T. Epplen, A. J. Jeffreys (Eds.). DNA fingerprinting: state of the science (pp. 211-219). Birkhäuser, Basilea: Springer.
Goicoechea, A. S., Carnese, F. R., Dejean, C., Avena, S. A., Weimer, T. A., Franco, M. H. L. P., Callegari-Jacques, S. M., Estalote, A. C., Simões, M. L. M. S., Palatnik, M., y Salzano, F. M. (2001). Genetic relationships between amerindian populations of Argentina. American Journal of Physical Anthropology , 115, 133-143.
Gómez-Carballa, A., Pardo-Seco, J., Brandini, S., Achilli, A., Perego, U. A., Coble, M. D., Diegoli, T. M., Álvarez-Iglesias, V., Martinón-Torres, F., Olivieri, A., Torroni, A., y Salas, A. (2018). The peopling of South America and the trans-Andean gene flow of the first settlers. Genome Research , 28, 767-779.
González Lebrero, R. (2002). La pequeña aldea. Sociedad y economía en Buenos Aires (1580-1640) Buenos Aires: Biblos.
Hofreiter, M., Paijmans, J., Goodchild, H., Speller, C., Barlow, A., Fortes, G., Thomas, J., Ludwig, A. y Collins, M. (2015). The future of ancient DNA: Technical advances and conceptual shifts. BioEssays, 37, 284-293.
Kashani, B. H., Perego, U. A., Olivieri, A., Angerhofer, N., Gandini, F., Carossa, V., Lancioni, H., Semino, O., Woodward, S. R., Achilli, A. y Torroni, A. (2012). Mitochondrial haplogroup C4c: a rare lineage entering America through the ice-free corridor? American Journal of Physical Anthropology , 147, 35-39.
Kozameh, L. y Brunás, O. (2013). Enfermedad de Paget en un individuo prehispánico del Delta del Paraná, confirmado por examen histológico y datación radiocarbónica. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano
-Series Especiales, 1(1), 114-120.
Lightfoot, K. (2005). Indians, missionaries and merchants. The legacy of colonial encounters on the California frontiers Los Ángeles: Editorial University of California Press.
Llamas, B., Fehren-Schmitz, L., Valverde, G., Soubrier, J., Mallick, S., Rohland, N., Nordenfelt, S., Valdiosera, C., Richards, S., Rohrlach, A., Barreto Romero, M., Flores Espinoza, I., Tomasto Cagigao, E., Watson Jiménez, L., Makowski, K., Leboreiro Reyna, I., Mansilla Lory, J., Ballivián Torrez, J., Rivera, M., Burger, R., Ceruti, C., Reinhard, J., Wells, R., Politis, G., Santoro, C., Standen, V., Smith, C., Reich, D., Ho, S., Cooper, A. y Haak, W. (2016). Ancient mitocondrial DNA provides high-resolution time scale of the peopling of the Americas. Science Advances, 2: e1501385. doi: 10.1126/sciadv.1501385
Lopes de Souza, P. (1969 [1531]). Diario de Navegação de Pero Lopes de Sousa (de 1530 a 1532). Revista Trimensal do Instituto Historio Geographico e Etnographico do Brasil, 24, 9-74.
Loponte, D. (2008). Arqueología del humedal del Paraná inferior (Bajíos Ribereños Meridionales) Buenos Aires: Instituto Nacional de Antropología y Pensamiento Latinoamericano.
Loponte, D., Acosta, A., y Musali, J. (2004). Complejidad social: cazadores-recolectores y horticultores en la región pampeana. En Martínez, G., Gutiérrez, M. A., Curtoni, R., Berón, M. y Madrid, P. (Eds.). Aproximaciones contemporáneas a la arqueología pampeana: Perspectivas teóricas, metodológicas, analíticas y casos de estudio (pp. 41-60). Buenos Aires: Universidad Nacional del Centro de la Provincia de Buenos Aires, Argentina.
Lothrop, S. (1932). Indians of the Paraná Delta. Annals of the New York of Sciences, 33, 77-232.
Marrero, A., Silva-Junior, W., Bravi, C., Hutz, M., Petzl-Erler, M., Ruiz-Linares, A., Salzano, F. y Bortolini, M. (2007). Demographic and evolutionary trajectories of the Guarani and Kaingang natives of Brazil. American Journal of Physical Anthropology , 132, 301-310.
Mazza, B. y Loponte, D. (2012). Las prácticas mortuorias en el humedal del Paraná inferior. Arqueología Iberoamericana, 13, 3-21.
Melo dos Santos, E., Soares da Silva, A., Daguer Ewerton, P., Takeshita, L., Thomaz Maia, M. (2015). Origins and demographic dynamics of Tupi expansion: a genetic tale. Boletim do Museu Paraense Emilio Goeldi. Ciências Humanas, 10, 217-228.
Motti, J. M. B., Muzzio, M., Ramallo, V., García, A., Alfaro, E. L., Dipierri, J. E., Bailliet, G., Coble, M. y Bravi, C. (2009). No es lo que parece: sitios diagnóstico revertantes en el ADN mitocondrial. En Actas de las Novenas Jornadas Nacionales de Antropología Biológica (p. 147). Puerto Madryn, Argentina.
Nores, R. y Demarchi, D. (2011). Análisis de haplogrupos mitocondriales en restos humanos de sitios arqueológicos de la provincia de Córdoba. Revista Argentina de Antropología Biológica , 13, 43-54.
O’Fallon, B. y Fehren-Schmitz, L. (2011). Native Americans experienced a strong population bottleneck coincident with European contact. PNAS, 108, 1-5.
Pauro, M., García, A., Nores, R. y Demarchi, D. (2013). Analysis of uniparental lineages in two villages of Santiago del Estero, Argentina, seat of “Pueblo de Indios” in colonial times. Human Biology, 85, 699-719.
Perego, U. A., Achilli, A., Angerhofer, N., Accetturo, M., Pala, M., Olivieri, A., Kashani, B. H., Ritchie, K. H., Scozzari, R., Kong, O. P., Myres, N. M., Salas, A., Semino, O., Bandelt, H. J., Woodward, S. R. y Torroni, A. (2009). Distinctive Paleo-Indian migration routes from Beringia marked by two rare mtDNA haplogroups. Current Biology, 19(1), 1-8.
Perego, U., Angerhofer, N., Pala, M., Olivieri, A., Lancioni, H., Kashani, B. H., Carossa, V., Ekins, J. E., Gómez-Carballa, A., Huber, G., Zimmermann, B., Corach, D., Babudri, N., Panara, F., Myres, N. M., Parson, W., Semino, O., Salas, A., Woodward, S. R., Achilli, A. y Torroni, A. (2010). The initial peopling of the Americas: A growing number of founding mitochondrial genomes from Beringia. Genome Research , 20, 1174-1179.
Rizzo, F., Cardozo, D. y Tapia, A. (2016). Múltiples líneas de evidencia aplicadas al estudio de un individuo prehispánico, sitio Rancho José (Buenos Aires). Revista Argentina de Antropología Biológica , 18, 1-15.
Sala, A., Argüelles, C., Marino, M., Bobillo, C., Fenocchio, A. y Corach, D. (2010). Genetic analysis of six communities of Mbyá-Guaraní inhabiting north eastern Argentina by mean of nuclear and mitochondrial polymorphic markers. Human Biology, 82, 433-456.
Schmidl, U. (1903 [1567]). Viaje al Río de la Plata, 1534-1554 Traducido por S. A. Lafone Quevedo. Buenos Aires: Editorial Cabaut y Cía.
Servini, F., Yao, D., Lomartire, L., Barbieri, A., Vianello, D., Ferri, G., Moretti, E., Dasso, M., Garagnani, P., Pettener, D., Franceschi, C., Luiselli, D. y Francesch, Z. (2013). Analysis of Population Substructure in Two Sympatric Populations of Gran Chaco, Argentina. PLoS One, 8(5): e64054. doi:10.1371/journal.pone.0064054
Silva Noelli, F. (2004). La distribución geográfica de las evidencias arqueológicas guaraní. Revista de Indias, 64(230), 17-34.
Stone, A. y Stoneking, M. (1998). mtDNA analysis of a prehistoric Oneota population: implications for the peopling of the New World. American Journal of Human Genetics, 62, 1153-1170.
Tapia, A. (2002). Indicadores biológicos y culturales de la conquista en la desembocadura del Río Paraná (Siglos XVII y XVIII). Relaciones de la Sociedad Argentina de Antropología , 27, 357-374.
Tapia, A. (2014). Cambio cultural y persistencia de las identidades nativas en la sociedad colonial de Baradero (siglos XVII y XVIII). Revista Teoría y Práctica de la Arqueología Histórica Latinoamericana, 3, 43-59.
Tapia, A., Landa, C., De Rosa, H. y Montanari, E. (2009). Artefactos metálicos de las inhumaciones del Cementerio Indígena de Baradero. En Arqueometría Latinoamericana, 2do. Congreso Argentino, 1ero. Latinoamericano, Volumen I (pp. 263-269). Buenos Aires: Comisión Nacional de Energía Atómica.
Tapia, A., Néspolo, E. y Noya, V. (2015). Santiago del Baradero. Origen y Evolución de un Pueblo de Indios Buenos Aires: Editorial Pangea Ediciones de la Tierra.
Torres, L. (1911). Los primitivos habitantes del Delta del Paraná Buenos Aires: Coni Hermanos.
Torres, M., Bravi, C., Bortolini, M., Duque, C., Callegari-Jacques, S., Ortiz, D., Bedoya, G., Groot de Restrepo, H. y Ruiz-Linares, A. (2006). Revertan to the major founder native American haplogroup C common in populations from northern South America. American Journal of Human Biology, 18, 59-65.
Torroni, A., Schurr, T., Cabell, M., Brown, M., Neel, J., Larsen, M., Smith, D., Vullo, C. y Wallace, D. (1993). Asian affinities and continental radiation of the four founding Native American mtDNA´s. American Journal of Human Genetics , 53, 563-590.